Artikli meditsiiniline ekspert

Uued väljaanded
Munasarjad
Viimati vaadatud: 04.07.2025

Kõik iLive'i sisu vaadatakse meditsiiniliselt läbi või seda kontrollitakse, et tagada võimalikult suur faktiline täpsus.
Meil on ranged allhanke juhised ja link ainult mainekate meediakanalite, akadeemiliste teadusasutuste ja võimaluse korral meditsiiniliselt vastastikuste eksperthinnangutega. Pange tähele, et sulgudes ([1], [2] jne) olevad numbrid on nende uuringute linkideks.
Kui tunnete, et mõni meie sisu on ebatäpne, aegunud või muul viisil küsitav, valige see ja vajutage Ctrl + Enter.
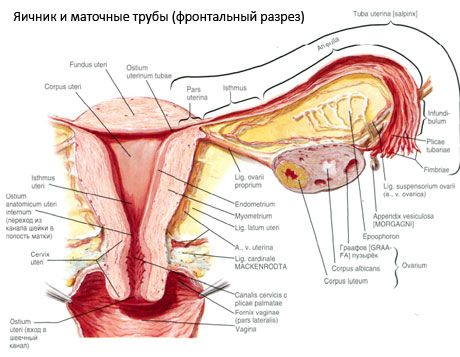
Munasari (ovarium; kreeka keeles oophoron) on paarisorgan, naissugunäärme, mis asub vaagnaõõnes emaka laia sideme taga. Munasarjades arenevad ja küpsevad naissugurakud (munarakud) ning moodustuvad naissuguhormoonid, mis sisenevad verre ja lümfi. Munasari on munaja kujuga, anteroposterior suunas mõnevõrra lamenenud. Munasarja värvus on roosakas. Sünnitanud naise munasarja pinnal on näha lohke ja arme - ovulatsiooni jälgi ja kollaskeha transformatsiooni. Munasari kaalub 5-8 g. Munasarja mõõtmed on: pikkus 2,5-5,5 cm, laius 1,5-3,0 cm, paksus - kuni 2 cm. Munasarjal on kaks vaba pinda: vaagnaõõne poole suunatud mediaalne pind (facies medialis), mis on osaliselt kaetud munajuhaga, ja külgmine pind (facies lateralis), mis külgneb vaagna külgseinaga kuni kergelt väljendunud lohuni - munasarja lohuni. See lohk asub nurgas, mis jääb ülalt kõhukelmega kaetud väliste niudeluu veresoonte ja alt emaka- ning sulgurarterite vahele. Munasarja taga kulgeb vastava poole kusejuha retroperitoneaalselt ülevalt alla.
Munasarja pinnad lähevad üle kumerasse vabasse (tagumisse) serva (margo liber), ees - mesenteerilisse serva (margo mesovaricus), mis on lühikese kõhukelme (munasarja mesenteeriumi) voldi abil kinnitunud emaka laia sideme tagumise lehekese külge. Sellel elundi esiserval on sooneline lohk - munasarja hilum (hilum ovarii), mille kaudu sisenevad munasarja arter ja närvid ning väljuvad veenid ja lümfisooned. Munasarjal on ka kaks otsa: ümar ülemine torukujuline ots (extremitas tubaria), mis on suunatud munajuha poole, ja alumine emakaots (extremitas utenna), mis on emakaga ühendatud munasarja sideme (lig. ovarii proprium) abil. See umbes 6 mm paksune ümmarguse nööri kujuline side kulgeb munasarja emakapoolsest otsast emaka külgnurgani, mis asub emaka laia sideme kahe lehe vahel. Munasarja siduaparaadi hulka kuulub ka munasarja sidukikinnitus (lig.suspensorium ovarii), mis on kõhukelme volt, mis kulgeb väikese vaagna seinast munasarjani ning sisaldab munasarja veresooni ja kiuliste kiudude kimpe. Munasarja kinnitab lühike mesenteer (mesovarium), mis on kõhukelme duplikaat, mis kulgeb emaka laia sideme tagumisest lehest munasarja mesenteerilise servani. Munasarjad ise ei ole kõhukelmega kaetud. Munajuha suurim munasarjaäär on kinnitunud munasarja torukujulise otsa külge. Munasarja topograafia sõltub emaka asendist ja suurusest (raseduse ajal). Munasarjad on väikese vaagnaõõne väga liikuvad organid.
 [
[ Munasarja veresooned ja närvid
Munasarjade verevarustus toimub aa. et vv. ovaricae et uterinae kaudu. Mõlemad munasarjaarterid (aa. ovaricae dextra et sinistra) algavad aordi esipinnalt neeruarterite alt; parempoolne pärineb tavaliselt aordist ja vasakpoolne neeruarterist. Suundudes allapoole ja külgsuunas mööda psoas major lihase esipinda, ületab iga munasarjaarter eesmise kusejuha (andes sellele harusid), välised niudeluu veresooned, äärejoone ja siseneb vaagnaõõnde, paiknedes siin munasarja suspensioonsidemes. Mediaalses suunas liikudes läbib munasarjaarter emaka laia sideme lehtede vahelt munajuha all, andes sellele harusid, ja seejärel munasarja mesenteeriumisse; see siseneb munasarja hilumisse.
Munasarjaarteri harud anastomoosivad laialdaselt emakaarteri munasarjaharudega. Munasarjade venoosne väljavool toimub peamiselt munasarjaveenide põimikusse, mis asub munasarja hilumis. Siit liigub veri kahes suunas: läbi emaka- ja munasarjaveenide. Paremal munasarjaveenil on klapid ja see suubub alumisse õõnesveeni. Vasak munasarjaveen suubub vasakusse neeruveeni, kuigi sellel puuduvad klapid.
Munasarjadest toimub lümfidrenaaž lümfisoonte kaudu, eriti rohkelt organivärava piirkonnas, kus eristub subovarianaalne lümfipõimik. Seejärel suundub lümf mööda munasarjade lümfisoonteid paraaortaalsetesse lümfisõlmedesse.
Munasarjade innervatsioon
Sümpaatiline - mida pakuvad tsöliaakia (päikese), ülemise mesenteerilise ja hüpogastrilise plexuse postganglionaarsed kiud; parasümpaatiline - vistseraalsete sakraalsete närvide poolt.
Munasarja struktuur
Munasarja pinda katab ühekihiline germinaalepiteel. Selle all asub tihe sidekoeline valgukest (tunica albuginea). Munasarja sidekude moodustab selle strooma (stroma ovarii), mis on rikas elastsete kiudude poolest. Munasarja aine ehk parenhüüm jaguneb välimiseks ja sisemiseks kihiks. Sisemist kihti, mis asub munasarja keskel, selle värava lähedal, nimetatakse medullaks (medulla ovarii). Selles kihis, lahtises sidekoes, on arvukalt vere- ja lümfisooni ning närve. Munasarja välimine kiht - koor (cortex ovarii) - on tihedam. See sisaldab palju sidekude, milles asuvad küpsevad primaarsed munasarja folliikulid (folliculi ovarici primarii), sekundaarsed (vesikulaarsed) folliikulid (folliculi ovarici secundarii, s.vesiculosi), samuti küpsed folliikulid, Graafi folliikulid (folliculi ovarici maturis), samuti kolla- ja atreetilised kehad.
Igas folliikulis on emaslooma munarakk ehk ootsüüt (ococytus). Munarakk on läbimõõduga kuni 150 µm, ümmargune, sisaldab tuuma ja suurel hulgal tsütoplasmat, mis lisaks rakuorganellidele sisaldab munaraku toitmiseks vajalikke valgu-lipiidide inklusioone (munakollane) ja glükogeeni. Munarakk kasutab oma toitainetevaru tavaliselt ära 12–24 tunni jooksul pärast ovulatsiooni. Kui viljastumist ei toimu, siis munarakk sureb.
Inimese munarakku katab kaks membraani. Sees on tsütolemma, mis on munaraku tsütoplasmaatiline membraan. Tsütolemma välisküljel on nn follikulaarsete rakkude kiht, mis kaitseb munarakku ja täidab hormoonide moodustamise funktsiooni – nad eritavad östrogeene.
Emaka, munajuhade ja munasarjade füsioloogilise asendi tagavad riputus-, kinnitus- ja tugiaparaadid, mis ühendavad kõhukelme, sidemeid ja vaagna kude. Riputusaparaat on esindatud paarismoodustistena, see hõlmab emaka ümar- ja laiasid sidemeid, munasarjade õigeid sidemeid ja riputussidemeid. Emaka laiad sidemed, munasarjade õiged ja riputussidemed hoiavad emakat keskmises asendis. Ümarad sidemed tõmbavad emaka põhja ettepoole ja tagavad selle füsioloogilise kalde.
Fikseerimis- (ankurdamis-)aparaat tagab emaka asukoha väikese vaagna keskel ja muudab praktiliselt võimatuks selle nihkumise külgedele, edasi-tagasi. Kuna aga sidemete aparaat eemaldub emakast alaosas, on võimalik emaka kaldumine erinevates suundades. Fikseerimisaparaat hõlmab sidemeid, mis asuvad vaagna lahtises koes ja ulatuvad emaka alaosast vaagna külg-, ees- ja tagaseinani: sakromaagilised, kardinaalsed, uterovesikaalsed ja vesikopubiaalsed sidemed.
Lisaks mesovariumile eristatakse järgmisi munasarjade sidemeid:
- Munasarja riputusside, mida varem nimetati infundibulopelviaalseks sidemeks. See on kõhukelme volt, mille sees läbivad veresooned (a. et v. ovarica) ja lümfisooned ning munasarja närvid, mis on venitatud vaagna külgseina, nimmefastsia (ühise niudearteri väliseks ja sisemiseks jagunemise piirkonnas) ja munasarja ülemise (munajuhade) otsa vahele.
- Munasarja õige side läbib laia emakasideme lehtede vahelt, tagumise lehe lähemalt, ja ühendab munasarja alumist otsa emaka külgmise servaga. Munasarja õige side kinnitub emaka külge munajuha alguse ja ümarsideme vahel, viimase taga ja kohal. Sideme paksuses läbivad rr. ovarii, mis on emakaarteri lõppharud;
- pimesoole-munasarja side Side ulatub pimesoole mesenteeriumist paremasse munasarja või emaka laia sidemeni kõhukelme voldi kujul. Side on ebapüsiv ja seda täheldatakse 1/2 - 1/3 naistest.
Toetavat aparaati esindavad vaagnapõhja lihased ja fastsiad, mis on jagatud alumiseks, keskmiseks ja ülemiseks (sisemiseks) kihiks.
Kõige võimsam on ülemine (sisemine) lihaskiht, mida esindab päraku tõstev paarlihas. See koosneb lihaskimpudest, mis ulatuvad sabaluudest vaagna luudeni kolmes suunas (pubococcygeus, iliococcygeus ja ischiococcygeus). Seda lihaskihti nimetatakse ka vaagna diafragmaks.
Keskmine lihaskiht asub sümfüüsi, häbemeluu ja istmikuluu vahel. Keskmine lihaskiht - urogenitaalne diafragma - hõivab vaagna väljavooluava eesmise poole, mille kaudu läbivad kusiti ja tupp. Selle lehtede vahel eesmises osas asuvad lihaskimbud, mis moodustavad kusiti välise sulgurlihase, tagumises osas aga lihaskimbud, mis kulgevad ristisuunas - perineumi sügav põiklihas.
Vaagnapõhjalihaste alumine (välimine) kiht koosneb pindmistest lihastest, mille kuju meenutab numbrit 8. Nende hulka kuuluvad bulbokavernoosne, ischiokavernoosne, väline päraku sulgurlihas ja pindmine põiki perineaallihas.
Munasarjade ontogenees
Folliikulite kasvu ja atresia protsess algab 20. rasedusnädalal ja sünnituse ajaks on tüdruku munasarjades kuni 2 miljonit munarakku. Menarheks väheneb nende arv 300 tuhandeni. Kogu reproduktiivse eluea jooksul saavutab küpsuse ja ovulatsiooni mitte rohkem kui 500 folliikulit. Folliikulite esialgne kasv ei sõltu FSH stimulatsioonist, on piiratud ja atresia tekib kiiresti. Arvatakse, et steroidhormoonide asemel on primaarsete folliikulite kasvu ja atresia peamiseks regulaatoriks lokaalsed autokriinsed/parakriinsed peptiidid. Arvatakse, et folliikulite kasvu ja atresia protsessi ei katkesta ükski füsioloogiline protsess. See protsess jätkub igas vanuses, sealhulgas emakasisese perioodi ja menopausi ajal, ning selle katkestavad rasedus, ovulatsioon ja anovulatsioon. Mehhanism, mis käivitab folliikulite kasvu ja nende arvu igas konkreetses tsüklis, on siiani ebaselge.
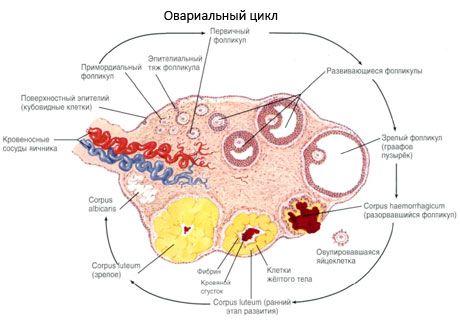
Folliikul läbib oma arengu käigus mitu etappi. Primaarsed idurakud pärinevad munakollase endodermist, allantoisist ja rändavad embrüo suguelundite piirkonda 5.-6. rasedusnädalal. Kiire mitootilise jagunemise tulemusena, mis jätkub 6.-8. rasedusnädalast kuni 16.-20. rasedusnädalani, moodustub embrüo munasarjades kuni 6-7 miljonit munarakku, mida ümbritseb õhuke granuloosrakkude kiht.
Preantraalne folliikul - munarakku ümbritseb membraan (Zona pellucida). Munarakku ümbritsevad granulosarakud hakkavad vohama, nende kasv sõltub gonadotropiinidest ja korreleerub östrogeenide tasemega. Granulosarakud on FSH sihtmärgiks. Preantraalse folliikuli staadiumis on granulosarakud võimelised sünteesima kolme klassi steroide: indutseerivad peamiselt aromataasi aktiivsust, mis on peamine ensüüm, mis muundab androgeenid östradiooliks. Arvatakse, et östradiool on võimeline suurendama oma retseptorite arvu, avaldades granulosarakkudele otsest mitogeenset toimet, mis ei sõltu FSH-st. Seda peetakse parakriinseks faktoriks, mis võimendab FSH toimet, sealhulgas aromatiseerumisprotsesside aktiveerimist.
FSH retseptorid ilmuvad granuloosrakkude membraanidele kohe, kui folliikuli kasv algab. FSH vähenemine või suurenemine viib selle retseptorite arvu muutumiseni. Seda FSH toimet moduleerivad kasvufaktorid. FSH toimib G-valgu, adenülaattsüklaasi süsteemi kaudu, kuigi steroidogeneesi folliikulis reguleerib peamiselt FSH, on selles protsessis kaasatud palju tegureid: ioonkanalid, türosiinkinaasi retseptorid, sekundaarsete virgatsainete fosfolipaasisüsteem.
Androgeenide roll folliikulite varajases arengus on keeruline. Granulosa rakkudel on androgeeniretseptorid. Need ei ole mitte ainult FSH-indutseeritud östrogeenideks aromatiseerumise substraadid, vaid võivad madalatel kontsentratsioonidel aromatiseerumist ka suurendada. Kui androgeeni tase tõuseb, valivad preantraalsed granulosa rakud eelistatavalt mitte östrogeenideks aromatiseerumise raja, vaid lihtsama androgeeni raja 5a-reduktaasi kaudu, et muundada need androgeeniks, mida ei saa östrogeeniks muundada, pärssides seeläbi aromataasi aktiivsust. See protsess pärsib ka FSH ja LH retseptori moodustumist, peatades seeläbi folliikulite arengu.
Aromatiseerumisprotsessi käigus läbib kõrge androgeenitasemega folliikulid atresiaprotsessid. Folliikuli kasv ja areng sõltub selle võimest muuta androgeenid östrogeenideks.
FSH juuresolekul on follikulaarvedeliku domineerivaks aineks östrogeenid. FSH puudumisel - androgeenid. LH puudub follikulaarvedelikus tavaliselt tsükli keskpaigani. Niipea kui LH tase tõuseb, väheneb granulosarakkude mitootiline aktiivsus, tekivad degeneratiivsed muutused ja androgeeni tase folliikulis suureneb. Steroidide tase follikulaarvedelikus on kõrgem kui plasmas ja peegeldab munasarjarakkude: granulosa- ja teekarakkude funktsionaalset aktiivsust. Kui FSH ainus sihtmärk on granulosarakud, siis LH-l on palju sihtmärke - teekarakud, strooma- ja luteiinirakud ning granulosarakud. Nii granulosa- kui ka teekarakkudel on võime steroidogeneesiks, kuid granulosarakkudes domineerib aromataasi aktiivsus.
Vastuseks LH-le toodavad theca-rakud androgeene, mis seejärel granulosarakkude poolt FSH-indutseeritud aromatiseerimise teel östrogeenideks muundatakse.
Folliikuli kasvades hakkavad theca rakud ekspresseerima LH retseptorite, P450 sec ja 3beta-hüdroksüsteroiddehüdrogenaasi geene, insuliinilaadne kasvufaktor (IGF-1) suurendab sünergiliselt LH-ga geenide ekspressiooni, kuid ei stimuleeri steroidogeneesi.
Munasarjade steroidogenees on alati LH-sõltuv. Folliikuli kasvades ekspresseerivad theca-rakud ensüümi P450c17, mis moodustab kolesteroolist androgeeni. Granulosa-rakkudel seda ensüümi ei ole ja nad sõltuvad theca-rakkudest, et toota androgeenidest östrogeene. Erinevalt steroidogeneesist on follikulogenees FSH-sõltuv. Folliikuli kasvades ja östrogeeni taseme tõustes aktiveerub tagasisidemehhanism - FSH tootmine on pärsitud, mis omakorda viib folliikuli aromataasi aktiivsuse vähenemiseni ja lõpuks follikulaarse atreesiani apoptoosi (programmeeritud rakusurma) kaudu.
Östrogeenide ja FSH tagasisidemehhanism pärsib juba kasvama hakanud folliikulite arengut, kuid mitte dominantfolliikulit. Dominantfolliikulis on rohkem FSH retseptoreid, mis toetavad granuloosrakkude proliferatsiooni ja androgeenide aromatiseerumist östrogeenideks. Lisaks toimivad parakriinsed ja autokriinsed rajad antraalfolliikulite arengu olulise koordinaatorina.
Autokriinne/parakriinne regulaator koosneb peptiididest (inhibiin, aktivin, follistatiin), mida sünteesivad granulosa rakud vastusena FSH-le ja mis sisenevad follikulaarsesse vedelikku. Inhibiin vähendab FSH sekretsiooni; aktivin stimuleerib FSH vabanemist hüpofüüsist ja võimendab FSH toimet munasarjas; follistatiin pärsib FSH aktiivsust, võimalik, et aktiviiniga seondudes. Pärast ovulatsiooni ja kollaskeha arengut on inhibiin LH kontrolli all.
Munasarjarakkude kasvu ja diferentseerumist mõjutavad insuliinilaadsed kasvufaktorid (IGE). IGF-1 toimib granuloosrakkudele, põhjustades tsüklilise adenosiinmonofosfaadi (cAMP), progesterooni, oksütotsiini, proteoglükaani ja inhibiini taseme tõusu.
IGF-1 toimib theca rakkudele, põhjustades androgeenide tootmise suurenemist. Theca rakud omakorda toodavad tuumorinekroosifaktorit (TNF) ja epidermise kasvufaktorit (EGF), mida samuti reguleerib FSH.
EGF stimuleerib granuloosrakkude proliferatsiooni. IGF-2 on follikulaarse vedeliku peamine kasvufaktor ning selles leidub ka IGF-1, TNF-α, TNF-3 ja EGF-i.
Munasarjade funktsiooni parakriinse ja/või autokriinse regulatsiooni häired näivad mängivat rolli ovulatsioonihäiretes ja polütsüstiliste munasarjade tekkes.
Antraalse folliikuli kasvades suureneb folliikulite vedelikus östrogeeni sisaldus. Nende suurenemise haripunktis ilmuvad granuloosrakkudele LH retseptorid, toimub granuloosrakkude luteiniseerumine ja progesterooni tootmine suureneb. Seega preovulatoorsel perioodil põhjustab östrogeeni tootmise suurenemine LH retseptorite ilmnemise, LH omakorda põhjustab granuloosrakkude luteiniseerumist ja progesterooni tootmist. Progesterooni suurenemine vähendab östrogeeni taset, mis ilmselt põhjustab FSH teise piigi tsükli keskel.
Arvatakse, et ovulatsioon toimub 10–12 tundi pärast LH piiki ja 24–36 tundi pärast östradiooli piiki. Arvatakse, et LH stimuleerib munaraku redutseerumist ehk jagunemist, granuloosrakkude luteiniseerumist ning progesterooni ja prostaglandiini sünteesi folliikulis.
Progesteroon suurendab proteolüütiliste ensüümide aktiivsust, mis koos prostaglandiiniga osalevad folliikuli seina purunemises. Progesterooni poolt indutseeritud FSH piik võimaldab munarakul folliikulist väljuda, muutes plasminogeeni proteolüütiliseks ensüümiks plasmiiniks, ning tagab piisava hulga LH-retseptoreid luteaalfaasi normaalseks arenguks.
3 päeva jooksul pärast ovulatsiooni suurenevad granuloosrakud ja neisse ilmuvad iseloomulikud vakuoolid, mis on täidetud pigmendi, luteiiniga. Teka-luteaalrakud eristuvad teekast ja stroomast ning saavad osaks kollakehast. Kollaskehasse tungivad kapillaarid arenevad angiogeneesi faktorite mõjul väga kiiresti ning paranenud vaskularisatsiooniga suureneb progesterooni ja östrogeenide tootmine. Steroidogeneesi aktiivsust ja kollaskeha eluiga määrab LH tase. Kollaskeha ei ole homogeenne rakuline moodustis. Lisaks kahele kollaskeha tüübile sisaldab see endoteelirakke, makrofaage, fibroblaste jne. Suured kollaskeharakud toodavad peptiide (relaksiini, oksütotsiini) ja on steroidogeneesis aktiivsemad, omades suuremat aromataasi aktiivsust ja suuremat progesterooni sünteesi kui väikesed rakud.
Progesterooni tipp saabub 8. päeval pärast LH tippu. On täheldatud, et progesterooni ja östradiooli sekreteeritakse luteaalfaasis episoodiliselt korrelatsioonis LH pulseeriva väljundiga. Kollaskeha moodustumisega läheb inhibiini tootmise kontroll FSH-lt LH-le üle. Inhibiini tase suureneb koos östradiooli taseme tõusuga enne LH tippu ja jätkab suurenemist ka pärast LH tippu, kuigi östrogeeni tase langeb. Kuigi inhibiini ja östradiooli sekreteerivad granulosarakud, reguleerivad neid erinevad rajad. Inhibiini langus luteaalfaasi lõpus aitab kaasa FSH suurenemisele järgmise tsükli jaoks.
Kollaskeha väheneb väga kiiresti - 9.-11. päeval pärast ovulatsiooni.
Degeneratsiooni mehhanism on ebaselge ja ei ole seotud östrogeenide luteolüütilise rolli ega retseptoritega seotud mehhanismiga, nagu on täheldatud endomeetriumis. Kollaskeha poolt toodetavate östrogeenide rollile on veel üks seletus. On teada, et östrogeene on vaja progesterooni retseptorite sünteesiks endomeetriumis. Luteaalfaasi östrogeenid on tõenäoliselt vajalikud progesterooniga seotud muutusteks endomeetriumis pärast ovulatsiooni. Progesterooni retseptorite ebapiisav areng ebapiisava östrogeeni taseme tagajärjel on võimalik lisamehhanism viljatuse ja varajaste raseduse katkemiste tekkeks, mis on veel üks luteaalfaasi puudulikkuse vorm. Arvatakse, et kollaskeha eluiga kujuneb välja ovulatsiooni ajal. Ja see kindlasti taandub, kui seda raseduse tõttu inimese kooriongonadotropiin ei toeta. Seega viib kollaskeha taandareng östradiooli, progesterooni ja inhibiini taseme languseni. Inhibiini vähenemine kõrvaldab selle pärssiva toime FSH-le; Östradiooli ja progesterooni vähenemine võimaldab GnRH sekretsiooni väga kiiresti taastuda ja tagasisidemehhanismi eemaldamist hüpofüüsist. Inhibiini ja östradiooli vähenemine koos GnRH suurenemisega annab FSH-le tõuke LH asemel. FSH suurenemine viib folliikulite kasvuni koos järgneva domineeriva folliikuli valikuga ja uus tsükkel algab, kui rasedust ei toimu. Steroidhormoonidel on reproduktiivbioloogias ja üldises füsioloogias juhtiv roll. Need määravad inimese fenotüübi, mõjutavad südame-veresoonkonda, luude ainevahetust, nahka, üldist heaolu ja mängivad võtmerolli raseduse ajal. Steroidhormoonide toime peegeldab rakusiseseid ja geneetilisi mehhanisme, mis on vajalikud rakuvälise signaali edastamiseks rakutuuma, et tekitada füsioloogiline reaktsioon.
Östrogeenid tungivad difuusselt läbi rakumembraani ja seonduvad rakutuumas asuvate retseptoritega. Seejärel seondub retseptor-steroidkompleks DNA-ga. Sihtrakkudes viivad need interaktsioonid geenide ekspressioonini, valkude sünteesini ning spetsiifiliste rakkude ja kudede funktsioonideni.