
Munarakk
Viimati vaadatud: 23.04.2024

Kõik iLive'i sisu vaadatakse meditsiiniliselt läbi või seda kontrollitakse, et tagada võimalikult suur faktiline täpsus.
Meil on ranged allhanke juhised ja link ainult mainekate meediakanalite, akadeemiliste teadusasutuste ja võimaluse korral meditsiiniliselt vastastikuste eksperthinnangutega. Pange tähele, et sulgudes ([1], [2] jne) olevad numbrid on nende uuringute linkideks.
Kui tunnete, et mõni meie sisu on ebatäpne, aegunud või muul viisil küsitav, valige see ja vajutage Ctrl + Enter.
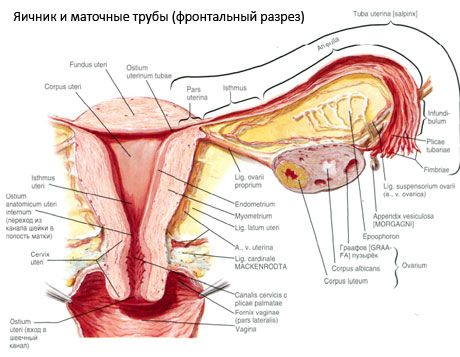
Munasarja (ovaarium, Kreeka oophoron) - ühendatud elund, naiste suguelundkond, mis asub väikese vaagna õõnsuses emaka laia sideme taga. Munasarjad arendavad ja küpsed naissoost sugurakud (ovules), samuti naissoost soohormoonid, mis sisenevad verd ja lümfi. Munasarjas on munakujuline kuju, mis on mõnevõrra lamestatud eesmise tagumise suunas. Munasarja värvus on roosakas. Sünnitava naise munaraku pinnal on nähtavad depressioonid ja armid - kollase keha ovulatsiooni jäljed ja ümberkujundamine. Munasarja mass on 5-8 g. Munasarjade mõõtmed on 2,5-5,5 cm pikad, 1,5-3,0 cm laiad ja 2 cm paksused. Munasarjal on kaks vaba pinda: facial medialis ) poole pööratud vaagnaõõs, osaliselt kattunud munajuha ja külgpinda (faatsieste lateralis), mis külgneb vaagna külgseinte, kerge süvendamist - munasarjade lohk. See fovea asub nurgas ülaosas olevate silma peritoneaalsete silmapiirkonna veresoonte ja emaka ja oklusuaalsete arterite vahel põhjas. Munasarja taga läbib vastav külg ülaosast allapoole.
Munasarja pind liigub kumer vaba (tagumine) serv (margo liber), ees - mesenteriaalsetes regioon (margo mesovaricus) abil lühiklipi-kordne kõhukelme (soolekinnistist munasari) tagaküljele lehtede kohta laisidemes emakas. Esirinnas organism ei rihveldatud süvend - munasarja paisu (seemnenaba ovarii), mille kaudu munasarjast hõlmavad artereid ja närve paiknev veenides ja lümfisoonte. Me isoleeriti ka munasarja kaks otsa: ümar ülemise torukujulise otsa (extremitas tubaria), ees munajuhas ja alumise otsa ema (extremitas utenna), seostati emakas enda kobaras munasarja (lig ovarii enda.). See kimp ümara juhtmest umbes 6 mm emalt on lõpuks külgmiste nurgas munasarjade emaka, vahel paiknevad kaks kihti laisidemes. By ligamentide aparaadi munasarja käsitleb ka ligament podveshivayaschaya munasarja (lig.suspensorium ovarii), mis on lahtikäiv kõhukelme ulatub ülemise seina vaagna munasarjakahjustust ja munasarjade mahuteid ja kiuline kiudude kimbud. Munarakkudes fikseeritud lühikese soolekinnistist (mesovarium), kelle duplikatury kõhukelme, mis ulatub tagumisse infolehte laisidemes emaka munasarjakahjustust mesenteersete serv. Munasarjad ise ei ole kõhukelmega kaetud. Emaka toru suurim munasarja fimbria on munarakkude otsa külge kinnitatud. Munasarja topograafia sõltub emaka asendist, selle suurusest (raseduse ajal). Munasarjad viitavad vaagnaõõne väga liikuvatele elunditele.
 [
[Munasarjad ja närvid
Munasarjade verevarustus tuleneb aa. Et vv. Ovaricae et uterinae. Mõlemad Munasarjaarter (. Aa ovaricae Dextra et sinistra) ulatuvad esipinna aordis veidi alla neeruarterid, paremal pärineb kõige sagedamini aorti ja vasakut - neeru arteri. Lähen alla ja külgsuunas üle esikülje nimmelihase, Munasarjaarter lõikub iga ees kusejuha (andes talle oksi), välise niude laevade ja piirijoone tulnud vaagnaõõne, paikneb siin peatava sideme munasarjas. Pärast medially, Munasarjaarter möödub linade vahel on laisidemes emaka all munajuhad, andes oksad ja siis - soolekinnistini munasarjast; siseneb munasarjade väravatele.
Munasarjade arterite oksad on laialdaselt anamneesiga emakaarteri munasarja okstega. Verejooks munasarjadest toimub peamiselt munasarjade väravas asuvas munasarjade venoosne plexus. Seega langeb vere väljavool kahes suunas: emaka ja munasarjaveenide kaudu. Õigus munasarja veenil on klapid ja jookseb alamvina kaavi. Vasakuline munasarja veen voolab vasakusse neeruvõõsisse, ilma ventiilideta.
Munasarjade lümfi väljavool tekib läbi lümfisõlmede, eriti rikkalikult orgaaniliste väravate piirkonnas, kus keeleline lümfipõimed on isoleeritud. Seejärel suunatakse lümfi munasarja lümfisõlmede para-aordi lümfisõlmedesse.
Munasarjade innervatsioon
Somaatiline - seda pakub tsöliaakia (päikese), ülemiste punaste ja hüpogastriapelkade postganglionide kiud; parasümpaatiline - sisemise sakraalne närvide tõttu.
Munasarja struktuur
Pind on kaetud ühekihiline munasarjade suguvõimeliste epiteeli. Selle all peitub tiheda sidekoe albuginea (tunica albuginea). Sidekoe munasarja moodustab strooma (stroomas ovarii), rikas elastne kiud. Aine munasarjade parenhüümi jaguneb see välimine kui sisemine kiht. Sisemine kiht, mis asub kesklinnas, munasarja lähemale paisu, nimetatakse medulla (medulla ovarii). Selles kihis lahtised sidekoe on arvukalt vere- ja lümfisoonte ja närve. Munasarja välimine kiht - koorikkollane munarakk on tihedam. See on palju sidekoe milles asuvad laagerdamine primaarse munasarjafolliikulid (folliculi ovarici primarii), sekundaarne (mulli) folliikulite (folliculi ovarici secundarii, s.vesiculosi), hästi küpse folliikulite graafovy mullid (folliculi ovarici maturis) ja kollane ja atretikehad.
Iga folliikuli puhul on naissoost reproduktiivne munarakk või munarakk (ovotsüte). Munarakku läbimõõduga kuni 150 mikronit, ümarad, sisaldab tuumas suurel hulgal tsütoplasmas mis lisaks rakulise organellid on Valkude ja lipiidide kandmisel (munarebu), glükogeeni vaja muna tarnimiseks. Tema ootsüütide tarbimine kulub tavaliselt 12-24 tundi pärast ovulatsiooni. Kui väetamist ei toimu, sureb muna.
Inimese munasarjal on kaks kattekihti. Sees on tsütomeem, mis on munarakkude tsütoplasmaatiline membraan. Väljaspool tsütomeemat on olemas nn follikulaarrakkude kiht, mis kaitsevad muna ja teostavad hormooni moodustavat funktsiooni - nad vabastavad östrogeene.
Emaka, torude ja munasarjade füsioloogilist positsiooni tagab vedeldav, fikseeriv ja toetav aparaat, kõhuõõne, sidemete ja vaagnapõhja tselluloos. Vedrustusseadet esindavad paarunud koosseisud, see hõlmab emaka ümmarguseid ja laiaid sidemeid, enda sidemeid ja libisevate munasarjade riputamist. Emaka suur sideme, ovaariumide omavahelist kinnihoidmist ja hoidmist emaka keskmises asendis. Ümarad sidemed meelitavad emaka põhja ettepoole ja tagavad selle füsioloogilise kalle.
Kinnitus (millega) seade annab nõrk positsioon keskel väike basseini ja on peaaegu võimatu kompenseerida osaliselt tagantjärele ja ettepoole. Kuid kuna sidumisseade liigub emaka alumisest osast eemal, on emakas võimalik kalduda mitmes suunas. Kinnitusvahenitega ühiku sisaldab nöörid paigutatud lahtise vaagna sidekoe ja ulatub alumise emaka kaardi küljele, esi- ja tagaseina vaagna: sacroiliac magochnye kardinal, emaka- ja tsüstiline-vesico-vaagnaliidese ligament.
Lisaks mesovariumile eristuvad järgmised munasarjade sidemed:
- peatav sideme munasarja varem määratud voronkotazovaya. On kordseid kõhukelme koos sees kulgeva vere (a. Et v. Ovarica) ja lümfisoonte ning närvide munasarja vahele tõmmatud külgne vaagnaseina, nimme sidekirme (jagamisel ühise niudearteritesse hetkel välimise ja sisemise) ja ülemiste (katseklaas) munasarja lõpp;
- munasarja enda sideme läbib lai emaka sideme lehti, tagumisele lehele lähemale ja ühendab munasarja alumise otsa koos emaka külgservaga. Emakule seotakse munasarja enda sideme emaka toru alguse ja ümmarguse sideme vahele, viimane neist edasi ja tagasi. Leiate paksuses on rr. Munarakk, mis on emakaarteri terminali harud;
- appendikulaarne-munasarjade sidumine Laiendab laiali parempoolse munarakkumi küljes oleva harja või emakakaela ulatusliku kõõlu kõhukelme vormi. Seos on ebastabiilne ja täheldatakse 1/2-1/3 naist.
Toestamisaparaati esindavad vaagnapõhja lihased ja fastsiaed, mis jagunevad alumiseks, keskmiseks ja ülemiseks (sisemiseks) kihiks.
Kõige võimsam on ülemine (sisemine) lihase kiht, mida esindab paarunud lihas, mis tõstab anni. Koosneb pikilihased, mille ventilaator välja õndraluu vaagna kolmes suunas (kubemekarvu-coccygeal, ilio-coccygeal ja ischiococcygeal lihased). Seda lihase kihti nimetatakse ka vaagna membraaniks.
Lihaste keskmine kiht asub sümfüüsi, kõri- ja ishhiumi luude vahel. Lihase keskmine kiht - urogenitaalmembraan - haarab vaagna väljavoolu esiosa, läbi selle läbib ureetra ja tupe. Vahel selle anterior infolehed on pikilihased mis moodustavad välise sulgurlihase kusiti, tagumisse ossa ette pikilihased sirutub välja ristisuunas, - deep põiki lahklihal lihasesse.
Alumine (välis) kiht vaagnapõhjalihaste koosneb pinna, mille asukoht on kujuline joonis 8. Nendeks bulbospongiosus-koopataoline, vaagnaliidese-koopataoline välise sulgurlihase päraku, pindmises põiki lahklihal lihasesse.
Munasarjade ontogenees
Kasvu- ja follikulaarse atresiooni protsess algab raseduse 20 nädala möödudes ja tüdruku munasarjade manustamise ajal jääb kuni 2 miljonit ootsüüti. Menstruatsiooni ajal väheneb nende arv 300 000-ni. Kogu reproduktiivterea jooksul saab küpsus ja ovulates mitte rohkem kui 500 folliikulit. Folliikulite esialgne kasv ei sõltu FSH-i stimuleerimisest, on piiratud ja atresia tekib kiiresti. Arvatakse, et steroidhormoonide asemel on kohalikud autokriinsed / parakriinpeptiidid peamine folliikulite kasvu ja atresia regulator. Usutakse, et füsioloogilised protsessid ei katkesta folliikulite kasvu ja atresia protsessi. See protsess jätkub igas vanuses, kaasa arvatud emakasisene periood ja menopaus, katkestatakse rasedus, ovulatsioon ja anovulatsioon. Folliikulite kasvu käivitav mehhanism ja nende arv igas konkreetses tsüklis ei ole veel selge.
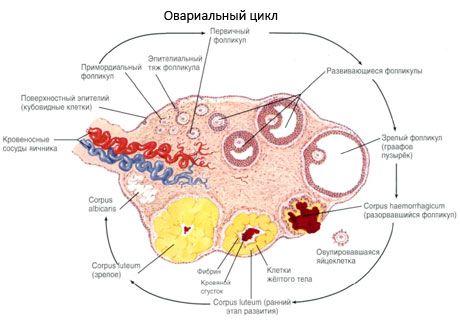
Selle arengus on folliikul läbi mitmeid arengutasemeid. Esmased idurakud pärinevad munarakkude endodermist, allantoiinist ja migreeruvad embrüo suguelundite piirkonda raseduse 5.-6. Nädala jooksul. Hepatiidi mitootilise jaotuse tulemusena, mis kestab 6-8 nädalat kuni 16-20 rasedusnädalani, moodustuvad embrüo munasarjades ümbritsetud granuloosrakkude õhuke kiht 6-7 miljonit ootsüüdit.
Preantaalfolliikli - munarakk ümbritseb membraani (Zona pellucida). Ootsüüte ümbritsevad granuloosrakud hakkavad vilkuma, nende kasv sõltub gonadotropiinidest ja korreleerub östrogeenide tasemega. FSH-i sihtmärgiks on granuloosrakud. Preantatiivse folliikuli staadiumis võivad granuloosrakud sünteesida kolme steroidide klassi: indutseerib eelistatavalt aromataasi, peamise ensüümi, mis muudab androgeenid östradiooliks, aktiivsust. Usutakse, et östradiool suudab suurendada oma retseptorite arvu, pakkudes otsest mitogeenset toimet FSH-st sõltumatutele granuloosrakkudele. Seda peetakse parakriinseks teguriks, mis suurendab FSH toimet, sealhulgas aromatiseerimise protsesside aktiveerimist.
FSH retseptorid ilmuvad granuloosrakkude membraanidele kohe, kui algab folliikulite kasv. FSH vähenemine või tõus viib tema retseptorite arvu muutumiseni. Seda FSH toimet moduleerivad kasvufaktorid. FSH toimib läbi G-valguga adenülaatsüntetaasi-tsüklaasi süsteemi folliiklites steroidogeneesile kuigi peamiselt reguleeritud FSH, hõlmab protsess paljudest teguritest: ioonkanalite retseptortürosiinkinaasi fosfolipaas süsteemi sekundaarse kullerid.
Androgeenide roll folliikulite varases arengus on keeruline. Granulosakrakkudel on androgeeni retseptorid. Need ei ole ainult substraati FSH indutseeritud aromatiseerimisest östrogeeni, kuid madalates kontsentratsioonides võib suurendada protsessi maitseaineid. Kui tase androgeenide suurendab preantral granuloosrakkudes eelistatavalt valitud tee no aromatiseerimisest viiakse östrogeenide ja lihtsam viis konverteerimise kaudu androgeenide 5a-reduktaasi arenevas androgeenide, mida ei saa muundada östrogeeni ja seeläbi pärsitud aromataasi aktiivsust. See protsess ka pärsib FSH ja LH retseptorite peatades sellega arengut folliikuli.
Aromatiseerimise protsess - kõrge asetusega androgeenide sisaldusega follikul läbib atresia protsesse. Folliikuli kasv ja areng sõltub tema võimest muuta androgeenid östrogeenideks.
FSH manulusel on follikulaarse vedeliku domineerivaks aineks östrogeenid. FSH-i puudumisel - androgeenid. LH on normaalne folliikulite vedelikus kuni tsükli keskpaigani. Niipea kui kõrgendatud mitootilise aktiivsuse LH granuloosrakkudes väheneb ja degeneratiivsed muutused tunduvad suurenenud androgeenide taseme folliiklites steroidi tasemete folliikulvedelik kui plasmas ning kajastab funktsionaalse aktiivsuse munasarja: granulosa ja teekarakkudel. Kui ainult sihtmärk FSH Granuloosrakud, siis LG on palju eesmärke - on teekarakkudel, stroomarakke ja kollaskeha ja granuloosrakkudes. Steroidogeneesi võime omab nii granuloosi kui ka teka rakke, kuid granulaosrakkudes domineerib aromataasi aktiivsus.
Vastuseks LH-ile tekitavad tecka rakud androgeene, mis seejärel FSH-ga indutseeritud aromatiseerimise teel transformeerivad granuloosrakud östrogeenidesse.
Nagu folliikuli teekarakkudel hakata väljendada geene LH retseptori P450 sek ja 3beeta-hüdroksüsteroiddehüdrogenaas, insuliini kasvufaktor (IGF-1) sünergiliselt LH suurendada geeniekspressioon, kuid ei stimuleeri steroidogeneesile.
Munasarjade steroidogenees on alati LH-sõltuv. Nagu folliikul kasvab, ekspresseerivad praegused rakud ensüümi P450c17, mis moodustab androgeeni kolesteroolist. Granuloosrakud ei ole selle ensüümi ja sõltuvad teekarakkudel on östrogeenide androgeenide. Erinevalt steroidogeneesist sõltub follikulogenees FSHist. Nagu folliikuli ja tõsta östrogeeni töölerakendumiseni tagasiside mehhanism - produktsiooni inhibeerimise FSH, mis omakorda toob kaasa languse aromataasi aktiivsust folliikuleid ning lõppkokkuvõttes folliikulisse atresia läbi apoptoosi (programmeeritud rakusurma).
Östrogeenide ja FSH-i tagasiside mehhanism pärsib kasvanud folliikulite arengut, kuid mitte domineerivat folliikuli. Domineeriv follikul sisaldab rohkem FSH retseptoreid, mis toetavad granuloosrakkude proliferatsiooni ja androgeenide aromatiseerimist östrogeenides. Lisaks sellele on parakriini- ja autokriinrajoon oluline antralfolliikuli arengu koordinaator.
Osa autokriini / parakriinne regulaator on peptiidid (inhibiin, aktiviin, follistatiin) granuloosrakud mis sünteesitakse vastuseks FSH ja saabuvad folliikulite vedelikku. Inhibiin vähendab FSH sekretsiooni; aktiviin stimuleerib FSH vabanemist hüpofüüsi ja suurendab FSH toimet munasarjas; Follitastiin pärsib FSH aktiivsust, mis on tingitud aktiini seondumisest. Pärast kollase keha ovulatsiooni ja arengut on inhibiin LH kontrolli all.
Munasarjade rakkude kasvu ja diferentseerumist mõjutavad insuliinitaolised kasvufaktorid (IGE). IGF-1 toimib granuloosrakkudes, põhjustades tsüklilise adenosiinmonofosfaadi (cAMP), progesterooni, oksütotsiini, proteoglükaani ja inhibiini suurenemist.
IGF-1 mõjutab teka rakke, põhjustades androgeenide produktsiooni suurenemist. Teka rakud toodavad omakorda kasvaja nekroosifaktorit (TNF) ja epidermaalset kasvufaktorit (EGF), mida reguleerivad ka FSH.
EGF stimuleerib granuloosrakkude proliferatsiooni. IGF-2 on follikulaarse vedeliku kasvu peamine tegur, see tuvastas ka IGF-1, TNF-a, TNF-3 ja EGF.
Näib, et parakriini ja / või autokriinset munasarjade funktsiooni reguleerimine on omakorda oluline ovulatsiooniprotsesside häirete ja polütsüstiliste munasarjade tekkimisel.
Kuna antralfolliikulis kasvab, suureneb folliikuliteedis sisalduvate östrogeenide sisaldus. Granuleosrakkude suurenemise tipul ilmnevad LH retseptorid, granuloosrakkude luteiniseerumine ja progesterooni tootmine suureneb. Seega, eelvulatoorsel perioodil põhjustab östrogeenide produktsiooni suurenemine LH retseptorite esilekutsumist, mis omakorda põhjustab granuloosrakkude luteiniseerumist ja progesterooni tootmist. Progesterooni suurenemine vähendab östrogeenide taset, mis ilmselt põhjustab FSH-i teise piigi tsükli keskel.
Usutakse, et ovulatsioon tekib 10 ... 12 tundi pärast LH piigi ja 24 ... 36 tundi pärast östradiooli piigi. Arvatakse, et LH stimuleerib munarakkude vähenemist, granuloosrakkude luteiniseerumist, progesterooni ja prostaglandiini sünteesi folliikulites.
Progesteroon suurendab proteolüütiliste ensüümide aktiivsust koos folliikulite seina purunemisega kaasatud prostaglandiiniga. FSH-indutseeritud progesterooni tipp, lahtrisse väljund munarakku folliikuleid muundades plasminogeeni proteolüütilise ensüümi - plasmiini annab piisava koguse LH retseptorid normaalseks arenguks luteaalfaasis.
3 päeva jooksul pärast ovulatsiooni granuloosrakud suureneb, need ilmuvad iseloomulikud vakuoolid täidetud pigmenti - luteiin. THECA-luteiini rakud diferentseeruvad alates teeka- ja strooma osaks kollaskeha. Väga kiiresti mõjul angiogeensed faktorid on kapillaaride arenemist levimist kollakeha ja parandamisega vaskularisatsioonita suurenenud progesterooni tootmist ja östrogeeni. Aktiivsus steroidogeneesile ja kestus kollakeha elu määrab LH tase. Kollane keha ei ole homogeenne rakuline üksus. Lisatud 2 tüüpi luteaalse rakud see sisaldab endoteeli rakud, makrofaagid, fibroblastid ja teised. Suured luteaalse rakud toodavad peptiidid (relaxin oksütotsiin) ja on aktiivsemad steroidogeneesile rohkem aromataasi aktiivsust ja suur progesterooni sünteesi kui väikestele rakkudes.
Progesterooni tipp on täheldatud kaheksandal päeval pärast LG tipphetkest. Märgiti, et progesteroon ja östradiool luteaalses faasis sekreteeritakse korrapäraselt korrelatsioonis LH-i impulsi väljundiga. Kollase keha moodustumisega kontrollib inhibiini tootmine FSH-st LH-i. Ingibin suureneb koos östradiooli suurenemisega LH-i tipuni ja suureneb pärast LH-i tippkoega, ehkki östrogeenide tase väheneb. Kuigi granuloosrakud sekreteerivad inhibiini ja östradiooli, reguleeritakse neid erinevalt. Inhibiini vähenemine luteali faasi lõpus suurendab FSH järgmisel tsüklil.
Kollane keha väga kiiresti - 9-11 päeva pärast ovulatsiooni väheneb.
Degeneratsiooni mehhanism ei ole selge ega ole seotud östrogeenide või retseptoriga seotud mehhanismi lüetootilise rolliga, nagu on näha endomeetriumis. Kollase keha tekitatud östrogeenide roll on veel üks selgitus. On teada, et progesterooni retseptorite sünteesiks endomeetriumis on vaja östrogeene. Luteaalse faasi östrogeenid on tõenäoliselt vajalikud progesterooniga seotud muutusteks endomeetriumil pärast ovulatsiooni. Ebapiisava östrogeeni sisalduse tõttu progesterooni retseptorite ebapiisav areng on ilmselt täiendav viljatuse ja varajase raseduse kaotuse mehhanism, mis on teine luteaalfaasi alanemise vorm. Arvatakse, et kollase keha eluiga on seatud ovulatsiooni ajal. Ja see sureb kindlasti tagasi, kui kooriongonadotropiini ei toetata seoses rasedusega. Seega põhjustab kollase keha regressioon östradiooli, progesterooni ja inhibiini taseme langust. Inhibiini vähendamine eemaldab selle inhibeeriva toime FSH-le; Östradiooli ja progesterooni vähendamine võimaldab väga kiiresti taastada GnRH sekretsiooni ja eemaldada hüpofüüsi tagasiside mehhanism. Inhibiini ja östradiooli vähendamine koos GnRH suurenemisega põhjustab FSH levimust LH suhtes. FSH-i suurenemine viib folliikulite kasvu, millele järgneb valitseva folliikuli valimine, ja algab uus tsükkel, kui rasedust ei toimu. Steroidhormoonid mängivad reproduktiivbioloogias ja üldiselt füsioloogias juhtivat rolli. Nad määravad inimese fenotüübi, mõjutavad kardiovaskulaarsüsteemi, luude, naha, keha üldist heaolu ja mängivad olulist rolli raseduse ajal. Steroidhormoonide toime peegeldab intratsellulaarset ja geneetilisi mehhanisme, mis on vajalikud rakuvälise signaali ülekandmiseks rakkude tuumale füsioloogilise vastuse esilekutsumiseks.
Estrogeenid hajuvad läbi rakumembraani ja seostuvad raku tuumas asuvate retseptoritega. Seejärel seondub retseptor-steroidkomplekt DNA-ga. Sihtrakkudes põhjustavad need koostoimed geeniekspressiooni, valkude sünteesi rakkude ja kudede spetsiifilisele funktsioonile.