Artikli meditsiiniline ekspert

Uued väljaanded
Katehhoolamiinide süntees, sekretsioon ja metabolism
Viimati vaadatud: 06.07.2025

Kõik iLive'i sisu vaadatakse meditsiiniliselt läbi või seda kontrollitakse, et tagada võimalikult suur faktiline täpsus.
Meil on ranged allhanke juhised ja link ainult mainekate meediakanalite, akadeemiliste teadusasutuste ja võimaluse korral meditsiiniliselt vastastikuste eksperthinnangutega. Pange tähele, et sulgudes ([1], [2] jne) olevad numbrid on nende uuringute linkideks.
Kui tunnete, et mõni meie sisu on ebatäpne, aegunud või muul viisil küsitav, valige see ja vajutage Ctrl + Enter.
Neerupealise säsi toodab ühendeid, mille struktuur on steroididest kaugel. Need sisaldavad 3,4-dihüdroksüfenüüli (katehhooli) tuuma ja neid nimetatakse katehhoolamiinideks. Nende hulka kuuluvad adrenaliin, noradrenaliin ja dopamiini beeta-hüdroksütüramiin.
Katehhoolamiinide sünteesi järjestus on üsna lihtne: türosiin → dihüdroksüfenüülalaniin (DOPA) → dopamiin → norepinefriin → adrenaliin. Türosiin siseneb organismi toiduga, kuid võib moodustuda ka fenüülalaniinist maksas fenüülalaniini hüdroksülaasi toimel. Türosiini muundamise lõppsaadused kudedes on erinevad. Neerupealise medullas jätkub protsess adrenaliini moodustumise staadiumis, sümpaatiliste närvide otstes - norepinefriini, mõnedes kesknärvisüsteemi neuronites lõpeb katehhoolamiinide süntees dopamiini moodustumisega.
Türosiini muundumist DOPA-ks katalüüsib türosiinhüdroksülaas, mille kofaktoriteks on tetrahüdrobiopteriin ja hapnik. Arvatakse, et see ensüüm piirab kogu katehhoolamiini biosünteesi protsessi kiirust ja seda pärsivad protsessi lõppsaadused. Türosiinhüdroksülaas on katehhoolamiini biosünteesi regulatiivsete mõjude peamine sihtmärk.
DOPA muundumist dopamiiniks katalüüsib ensüüm DOPA dekarboksülaas (kofaktor - püridoksaalfosfaat), mis on suhteliselt mittespetsiifiline ja dekarboksüleerib teisi aromaatseid L-aminohappeid. Siiski on märke võimalusest muuta katehhoolamiinide sünteesi selle ensüümi aktiivsust muutes. Mõnel neuronil puuduvad ensüümid dopamiini edasiseks muundamiseks ja see on lõpp-produkt. Teised koed sisaldavad dopamiini beeta-hüdroksülaasi (kofaktorid - vask, askorbiinhape ja hapnik), mis muundab dopamiini norepinefriiniks. Neerupealise medullas (kuid mitte sümpaatiliste närvide otstes) on fenüületanoolamiin - metüültransferaas, mis moodustab norepinefriinist adrenaliini. Sel juhul toimib S-adenosüülmetioniin metüülrühmade doonorina.
Oluline on meeles pidada, et fenüületanoolamiin-N-metüültransferaasi sünteesi indutseerivad glükokortikoidid, mis sisenevad neerupealise medullasse ajukoorest portaalveeni süsteemi kaudu. See võib selgitada asjaolu, et ühes organis on ühendatud kaks erinevat endokriinset nääret. Glükokortikoidide olulisust adrenaliini sünteesil rõhutab asjaolu, et noradrenaliini tootvad neerupealise medulla rakud paiknevad arteriaalsete veresoonte ümbruses, samas kui adrenaliini tootvad rakud saavad verd peamiselt neerupealise koores asuvatest venoossetest siinustest.
Katehhoolamiinide lagunemine toimub peamiselt kahe ensüümsüsteemi mõjul: katehhool-O-metüültransferaas (COMT) ja monoamiinoksüdaas (MAO). Adrenaliini ja noradrenaliini lagunemise peamised rajad on skemaatiliselt näidatud joonisel 54. COMT mõjul ja metüülrühma doonori S-adrenosüülmetioniini juuresolekul muunduvad katehhoolamiinid normetanefriiniks ja metanefriiniks (noradrenaliini ja adrenaliini 3-O-metüülderivaadid), mis MAO mõjul muunduvad aldehüüdideks ja seejärel (aldehüüdioksüdaasi juuresolekul) vanillüülmandelhappeks (VMA), mis on noradrenaliini ja adrenaliini peamine lagunemisprodukt. Samal juhul, kui katehhoolamiinid puutuvad esialgu kokku MAO-ga, mitte COMT-ga, muunduvad nad 3,4-dioksomandelaldehüüdiks ja seejärel aldehüüdioksüdaasi ja COMT mõjul 3,4-dioksomandelhappeks ja VMC-ks. Alkoholdehüdrogenaasi juuresolekul võib katehhoolamiinidest moodustuda 3-metoksü-4-oksüfenüülglükool, mis on KNS-is adrenaliini ja noradrenaliini lagunemise peamine lõpp-produkt.
Dopamiini lagunemine on sarnane, välja arvatud see, et selle metaboliitidel puudub beeta-süsiniku aatomi juures hüdroksüülrühm ja seetõttu moodustub vanillüülmandelhappe asemel homovanilliinhape (HVA) ehk 3-metoksü-4-hüdroksüfenüüläädikhape.
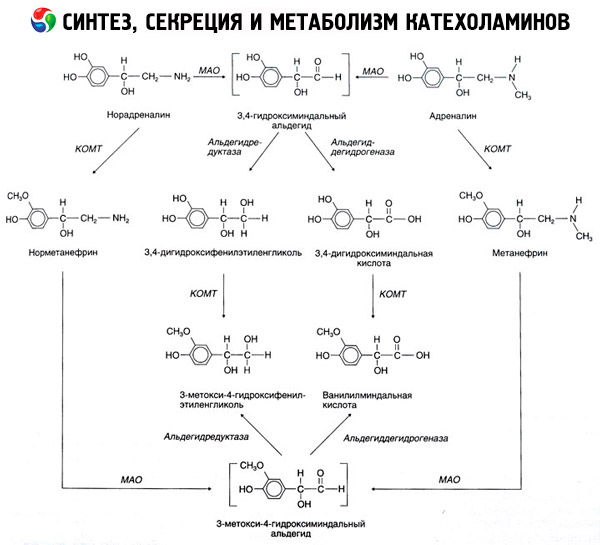
Samuti on oletatud katehhoolamiini molekuli kinoidse oksüdatsiooni raja olemasolu, mis võib toota väljendunud bioloogilise aktiivsusega vaheprodukte.
Sümpaatiliste närvilõpmete ja neerupealise säsi tsütosoolsete ensüümide toimel tekkinud norepinefriin ja adrenaliin sisenevad sekretoorsetesse graanulitesse, mis kaitseb neid lagundavate ensüümide toime eest. Katehhoolamiinide püüdmine graanulite poolt nõuab energiakulu. Neerupealise säsi kromafiingraanulites on katehhoolamiinid tihedalt seotud ATP-ga (vahekorras 4:1) ja spetsiifiliste valkudega - kromograniinidega, mis takistab hormoonide difusiooni graanulitest tsütoplasmasse.
Katehhoolamiinide sekretsiooni otsene stiimul on ilmselt kaltsiumi tungimine rakku, stimuleerides eksotsütoosi (graanulite membraani sulandumine rakupinnaga ja nende purunemine lahustuva sisu - katehhoolamiinide, dopamiini beeta-hüdroksülaasi, ATP ja kromograniinide - täieliku vabanemisega rakuvälisesse vedelikku).
Katehhoolamiinide füsioloogiline toime ja toimemehhanism
Katehhoolamiinide toime algab interaktsioonist sihtrakkude spetsiifiliste retseptoritega. Kui kilpnäärme- ja steroidhormoonide retseptorid paiknevad rakkude sees, siis katehhoolamiinide (samuti atsetüülkoliini ja peptiidhormoonide) retseptorid paiknevad raku välispinnal.
On juba ammu teada, et mõnede reaktsioonide puhul on adrenaliin või noradrenaliin efektiivsem kui sünteetiline katehhoolamiin isoproterenool, samas kui teiste puhul on isoproterenooli toime parem kui adrenaliinil või noradrenaliinil. Selle põhjal töötati välja kontseptsioon kahe tüüpi adrenoretseptorite esinemisest kudedes: alfa- ja beeta-adrenoretseptorid ning mõnes neist võib esineda ainult üks neist kahest tüübist. Isoproterenool on beeta-adrenoretseptorite kõige võimsam agonist, samas kui sünteetiline ühend fenüülefriin on alfa-adrenoretseptorite kõige võimsam agonist. Looduslikud katehhoolamiinid - adrenaliin ja noradrenaliin - on võimelised suhtlema mõlemat tüüpi retseptoritega, kuid adrenaliinil on suurem afiinsus beeta- ja noradrenaliinil alfa-retseptorite suhtes.
Katehhoolamiinid aktiveerivad südame beeta-adrenergilisi retseptoreid tugevamalt kui silelihaste beeta-retseptorid, mis võimaldas beeta-tüüpi jagada alatüüpideks: beeta1-retseptorid (süda, rasvarakud) ja beeta2-retseptorid (bronhid, veresooned jne). Isoproterenooli mõju beeta1-retseptoritele ületab adrenaliini ja noradrenaliini mõju vaid 10 korda, samas kui beeta2-retseptoritele toimib see 100–1000 korda tugevamalt kui looduslikud katehhoolamiinid.
Spetsiifiliste antagonistide (fentolamiin ja fenoksübensamiin alfa- ja propranolool beeta-retseptorite jaoks) kasutamine kinnitas adrenoretseptorite klassifikatsiooni adekvaatsust. Dopamiin on võimeline interakteeruma nii alfa- kui ka beeta-retseptoritega, kuid ka erinevatel kudedel (aju, hüpofüüs, veresooned) on oma dopamiinergilised retseptorid, mille spetsiifiline blokaator on haloperidool. Beeta-retseptorite arv varieerub 1000 kuni 2000 raku kohta. Beeta-retseptorite vahendatud katehhoolamiinide bioloogilised toimed on tavaliselt seotud adenülaattsüklaasi aktiveerimise ja cAMP rakusisese sisalduse suurenemisega. Kuigi retseptor ja ensüüm on funktsionaalselt seotud, on nad erinevad makromolekulid. Guanosiintrifosfaat (GTP) ja teised puriinnukleotiidid osalevad adenülaattsüklaasi aktiivsuse moduleerimises hormoon-retseptori kompleksi mõjul. Ensüümi aktiivsuse suurendamisega näivad nad vähendavat beeta-retseptorite afiinsust agonistide suhtes.
Denerveeritud struktuuride suurenenud tundlikkuse nähtus on juba ammu teada. Vastupidi, pikaajaline kokkupuude agonistidega vähendab sihtkudede tundlikkust. Beetaretseptorite uurimine on võimaldanud neid nähtusi selgitada. On näidatud, et pikaajaline kokkupuude isoproterenooliga viib adenülaattsüklaasi tundlikkuse kadumiseni beetaretseptorite arvu vähenemise tõttu.
Desensibiliseerumisprotsess ei nõua valgusünteesi aktiveerimist ja on tõenäoliselt tingitud pöördumatute hormoon-retseptori komplekside järkjärgulisest moodustumisest. Vastupidi, 6-oksüdopamiini sissetoomisega, mis hävitab sümpaatilisi lõpp-punkte, kaasneb reageerivate beeta-retseptorite arvu suurenemine kudedes. On võimalik, et sümpaatilise närvitegevuse suurenemine põhjustab ka vanusega seotud veresoonte ja rasvkoe desensibiliseerumist katehhoolamiinide suhtes.
Adrenoretseptorite arvu erinevates organites saab reguleerida teiste hormoonidega. Seega suurendab östradiool ja vähendab progesteroon alfa-adrenoretseptorite arvu emakas, millega kaasneb vastav selle kontraktiilse reaktsiooni suurenemine ja vähenemine katehhoolamiinidele. Kui beeta-retseptori agonistide toimel moodustunud rakusisene "teine sõnumitooja" on kindlasti cAMP, siis on olukord alfa-adrenergiliste efektide edastaja osas keerulisem. Eeldatakse mitmesuguste mehhanismide olemasolu: cAMP taseme langus, cAMP sisalduse suurenemine, rakulise kaltsiumidünaamika moduleerimine jne.
Erinevate toimete taasesitamiseks organismis on tavaliselt vaja adrenaliini annuseid, mis on 5-10 korda väiksemad kui noradrenaliinil. Kuigi viimane on α- ja β1-adrenergiliste retseptorite suhtes efektiivsem, on oluline meeles pidada, et mõlemad endogeensed katehhoolamiinid on võimelised interakteeruma nii alfa- kui ka beetaretseptoritega. Seetõttu sõltub antud organi bioloogiline reaktsioon adrenergilisele aktivatsioonile suuresti selles esinevate retseptorite tüübist. See aga ei tähenda, et sümpaatilise-neerupealise süsteemi närvi- või humoraalse sideme selektiivne aktivatsioon on võimatu. Enamasti täheldatakse selle erinevate sidemete suurenenud aktiivsust. Seega on üldtunnustatud, et hüpoglükeemia aktiveerib refleksiivselt neerupealise medulla, samas kui vererõhu langusega (posturaalne hüpotensioon) kaasneb peamiselt noradrenaliini vabanemine sümpaatiliste närvide otstest.
Adrenoretseptorid ja nende aktivatsiooni mõjud erinevates kudedes
Süsteem, organ |
Adrenergiliste retseptorite tüüp |
Reaktsioon |
Kardiovaskulaarsüsteem: |
||
Süda |
Beeta |
Suurenenud südame löögisagedus, juhtivus ja kontraktiilsus |
Arterioolid: |
||
Nahk ja limaskestad |
Alfa |
Vähendamine |
Skeletilihased |
Beeta |
Paisumine Kokkutõmbumine |
Kõhuõõne organid |
Alfa (rohkem) |
Vähendamine |
Beeta |
Laiendus |
|
Veenid |
Alfa |
Vähendamine |
Hingamissüsteem: |
||
Bronhide lihased |
Beeta |
Laiendus |
Seedesüsteem: |
||
Magu |
Beeta |
Vähenenud motoorsed oskused |
Sooled |
Alfa |
Sfinkterite kokkutõmbumine |
Põrn |
Alfa |
Vähendamine |
Beeta |
Lõõgastus |
|
Eksokriinne kõhunääre |
Alfa |
Vähenenud sekretsioon |
Kuseteede süsteem: |
Alfa |
Sulgurlihase kokkutõmbumine |
Põis |
Beeta |
Ejektorlihase lõdvestamine |
Meeste suguelundid |
Alfa |
Ejakulatsioon |
Silmad |
Alfa |
Pupillide laienemine |
Nahk |
Alfa |
Suurenenud higistamine |
Süljenäärmed |
Alfa |
Kaaliumi ja vee eritumine |
Beeta |
Amülaasi sekretsioon |
|
Endokriinsed näärmed: |
||
Kõhunäärme saarekesed |
||
Beetarakud |
Alfa (rohkem) |
Vähenenud insuliini sekretsioon |
Beeta |
Suurenenud insuliini sekretsioon |
|
Alfa-rakud |
Beeta |
Glükagooni sekretsiooni suurenemine |
8-rakuline |
Beeta |
Somatostatiini sekretsiooni suurenemine |
Hüpotalamus ja hüpofüüs: |
||
Somatotroofid |
Alfa |
STH sekretsiooni suurenemine |
Beeta |
STH sekretsiooni vähenemine |
|
Laktotroofid |
Alfa |
Prolaktiini sekretsiooni vähenemine |
Türeotroofid |
Alfa |
TSH sekretsiooni vähenemine |
Kortikotroofid |
Alfa |
Suurenenud AKTH sekretsioon |
| beeta | AKTH sekretsiooni vähenemine | |
Kilpnääre: |
||
Follikulaarsed rakud |
Alfa |
Türoksiini sekretsiooni vähenemine |
Beeta |
Türoksiini sekretsiooni suurenemine |
|
Parafollikulaarsed (K) rakud |
Beeta |
Kaltsitoniini sekretsiooni suurenemine |
Kõrvalkilpnäärmed |
Beeta |
PTH sekretsiooni suurenemine |
Neerud |
Beeta |
Suurenenud reniini sekretsioon |
Magu |
Beeta |
Suurenenud gastriini sekretsioon |
BX |
Beeta |
Suurenenud hapnikutarbimine |
Maks |
? |
Suurenenud glükogenolüüs ja glükoneogenees glükoosi vabanemisega; suurenenud ketogenees ketokehade vabanemisega |
Rasvkude |
Beeta |
Suurenenud lipolüüs vabade rasvhapete ja glütserooli vabanemisega |
Skeletilihased |
Beeta |
Suurenenud glükolüüs püruvaadi ja laktaadi vabanemisega; vähenenud proteolüüs alaniini ja glutamiini vabanemise vähenemisega |
Oluline on arvestada, et katehhoolamiinide intravenoosse manustamise tulemused ei kajasta alati adekvaatselt endogeensete ühendite toimet. See kehtib peamiselt norepinefriini kohta, kuna organismis ei vabane see peamiselt verre, vaid otse sünapsipiludesse. Seega aktiveerib endogeenne norepinefriin näiteks mitte ainult veresoonte alfaretseptoreid (vererõhu tõus), vaid ka südame beetareseptoreid (südame löögisageduse tõus), samas kui norepinefriini sissetoomine väljastpoolt viib peamiselt veresoonte alfaretseptorite aktiveerumiseni ja refleksiivse (vaguse kaudu) südame löögisageduse aeglustumiseni.
Adrenaliini väikesed annused aktiveerivad peamiselt lihasveresoonte ja südame beetaretseptoreid, mille tulemuseks on perifeerse veresoonte resistentsuse vähenemine ja südame väljundmahu suurenemine. Mõnel juhul võib esialgne efekt domineerida ja pärast adrenaliini manustamist tekib hüpotensioon. Suuremate annuste korral aktiveerib adrenaliin ka alfaretseptoreid, millega kaasneb perifeerse veresoonte resistentsuse suurenemine ja südame väljundmahu suurenemise taustal viib see arteriaalse rõhu tõusuni. Siiski säilib ka selle mõju veresoonte beetaretseptoritele. Selle tulemusena ületab süstoolse rõhu tõus sarnase näitaja diastoolse rõhu (pulsirõhu tõus). Veelgi suuremate annuste manustamisel hakkavad domineerima adrenaliini alfamimeetilised toimed: süstoolne ja diastoolne rõhk suurenevad paralleelselt, nagu norepinefriini mõjul.
Katehhoolamiinide mõju ainevahetusele koosneb nende otsestest ja kaudsetest efektidest. Esimesed realiseeruvad peamiselt beetaretseptorite kaudu. Keerulisemad protsessid on seotud maksaga. Kuigi maksa glükogenolüüsi suurenemist peetakse traditsiooniliselt beetaretseptori aktivatsiooni tulemuseks, on tõendeid ka alfaretseptorite osaluse kohta. Katehhoolamiinide kaudne mõju on seotud paljude teiste hormoonide, näiteks insuliini, sekretsiooni moduleerimisega. Adrenaliini mõjus selle sekretsioonile domineerib selgelt alfaadrenergiline komponent, kuna on näidatud, et iga stressiga kaasneb insuliini sekretsiooni pärssimine.
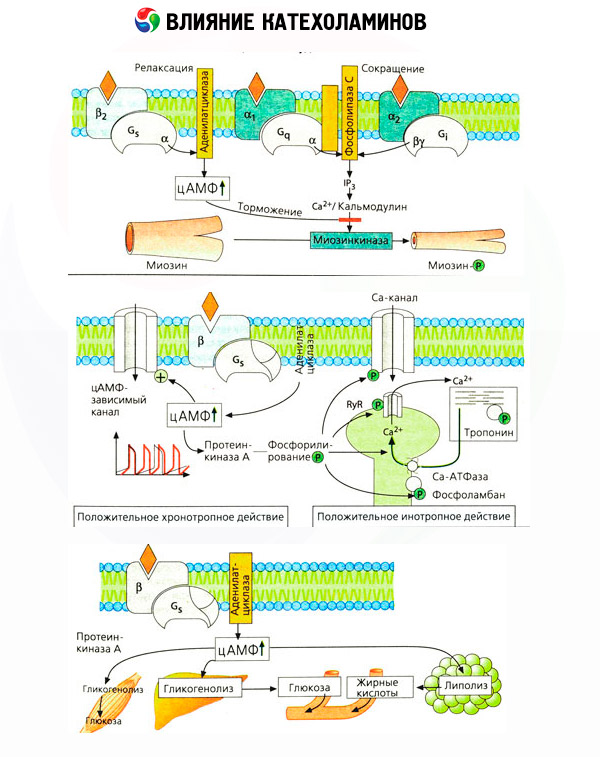
Katehhoolamiinide otsese ja kaudse toime kombinatsioon põhjustab hüperglükeemiat, mis on seotud mitte ainult suurenenud maksa glükoositootmisega, vaid ka selle kasutamise pärssimisega perifeersetes kudedes. Lipolüüsi kiirenemine põhjustab hüperlipatsideemiat koos rasvhapete suurenenud kohaletoimetamisega maksa ja ketokehade tootmise intensiivistumisega. Suurenenud glükolüüs lihastes viib laktaadi ja püruvaadi vabanemise suurenemiseni verre, mis koos rasvkoest vabaneva glütserooliga toimivad maksa glükoneogeneesi eelkäijatena.
Katehhoolamiinide sekretsiooni reguleerimine. Sümpaatilise närvisüsteemi ja neerupealise medulla produktide ja reaktsioonimeetodite sarnasus oli aluseks nende struktuuride ühendamisele keha üheks sümpaatilise-neerupealise süsteemiks, eraldades selle närvi- ja hormonaalsed seosed. Erinevad aferentsed signaalid on koondunud hüpotalamusesse ja seljaaju ning medulla oblongata keskustesse, kust pärinevad efferentsed sõnumid, mis lülituvad seljaaju külgmistes sarvedes asuvate preganglionaarsete neuronite rakulistele kehadele VIII emakakaela - II-III nimmepiirkonna segmentide tasandil.
Nende rakkude preganglionaarsed aksonid lahkuvad seljaajust ja moodustavad sünaptilisi ühendusi sümpaatilise ahela ganglionides asuvate neuronitega või neerupealise medulla rakkudega. Need preganglionaarsed kiud on kolinergilised. Esimene põhimõtteline erinevus sümpaatiliste postganglionaarsete neuronite ja neerupealise medulla kromafiinrakkude vahel on see, et viimased edastavad nende poolt vastuvõetud kolinergilist signaali mitte närvijuhtivuse (postganglionaarsed adrenergilised närvid), vaid humoraalse raja kaudu, vabastades adrenergilisi ühendeid verre. Teine erinevus on see, et postganglionaarsed närvid toodavad norepinefriini, neerupealise medulla rakud aga peamiselt adrenaliini. Neil kahel ainel on kudedele erinev mõju.