
Hüpofüüsi
Viimati vaadatud: 23.04.2024

Kõik iLive'i sisu vaadatakse meditsiiniliselt läbi või seda kontrollitakse, et tagada võimalikult suur faktiline täpsus.
Meil on ranged allhanke juhised ja link ainult mainekate meediakanalite, akadeemiliste teadusasutuste ja võimaluse korral meditsiiniliselt vastastikuste eksperthinnangutega. Pange tähele, et sulgudes ([1], [2] jne) olevad numbrid on nende uuringute linkideks.
Kui tunnete, et mõni meie sisu on ebatäpne, aegunud või muul viisil küsitav, valige see ja vajutage Ctrl + Enter.
Hüpofüüsi (hüpofüüs, s.glandula pituitaria) salvestatud ajuripatsi fossa sella kiilukujulise luu ja koljuõõnt eraldatakse tahke täiendit ajumembraani, moodustades diafragma iste. Selle diafragma ava kaudu on hüpofüüs ühendatud midbraani hüpotalamuse lehtriga. Hüpofüüsi põikisuurus on 10-17 mm, anteroposterior - 5-15 mm, vertikaalne - 5-10 mm. Hüpofüüsi mass meestel on umbes 0,5 g, naistel on 0,6 g. Väljas on hüpofüüsi kaetud kapsliga.
Vastavalt arengu ajuripatsi kahest erinevast mikroobe kehas eristada kahte ossa - ees ja taga. Adenohypophysis või varasem lobe (adenohypophysis, s.lobus anterior), seda suurem on 70-80% kogumassist ajuripatsis. See on tihedam kui tagumine laba. Eesmises lobe distaalse osa eraldatakse (pars distalis), millele kulub esiosa ajuripatsi lohk, vaheosa (pars intermedia), mis on paigutatud piirjoont tagumise jagage ja bugornuyu osa (pars tuberalis), jättes üles ja ühendatud lehtri hüpotaalamuse kasvajad. Veresoonte arvukuse tõttu on eesmine vähk kahvatukollane ja punakas toon. Parenhüümi hüpofüüsi eessagara näärmeliste esindajad mitut liiki rakke, mis asuvad harude vahel sinusoidi kapillaarid. Pool (50%) soovitud adenohypophysis rakud on hromafilnymi adenocytes võttes nende tsütoplasmas peengraanulite, hästi värviti kroomisoolade. See acidophilic adenocytes (40% adenohypophysis rakud) ja basofiilne adenocytes {10%). Arvu basofiili adenocytes hulka gonadotropic, kortikotropnye türeotroopne endocrinocytes. Kromofoobset adenocytes väike, neil on suur tuum ja väike kogus tsütoplasmas. Neid rakke peetakse kromofiilsete adenotsüütide prekursoriteks. Teised 50% adenohüpofüüsirakkudest on kromofobsed adenotsüüdid.
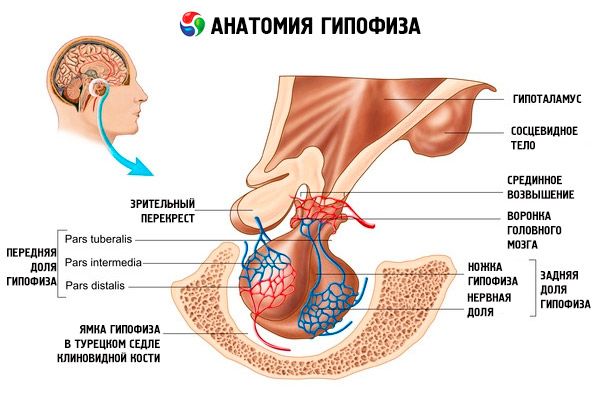
Neurohypophysis või posterior lobe (neurohypophysis, s.lobus posterior), kuhu kuuluvad närvi fraktsioon (Lõbus nervosus), mis asub tagaosas ajuripatsi lohk ja lehtri (infundibulum), mis asub taga bugornoy portsjoni adenohypophysis. Hüpofüüsi posterior lobe moodustatud gliiarakkude (ajuripatsi rakud), närvikiude ulatub neurosekretoorsete tuumades hüpotalamuse neurohypophysis ja neurosekretoorsete rakkudes.
Närvikiudude (rajad) ja veresoonte abil on hüpofüüsi funktsioon funktsionaalselt seotud aju hüpotalamusega, mis reguleerib hüpofüüsi aktiivsust. Hüpofüüsi ja hüpotalamuse koos nende neuroendokriinsuse, vaskulaarsete ja närviühendustega peetakse tavaliselt hüpotaalamuse-hüpofüüsi süsteemi.
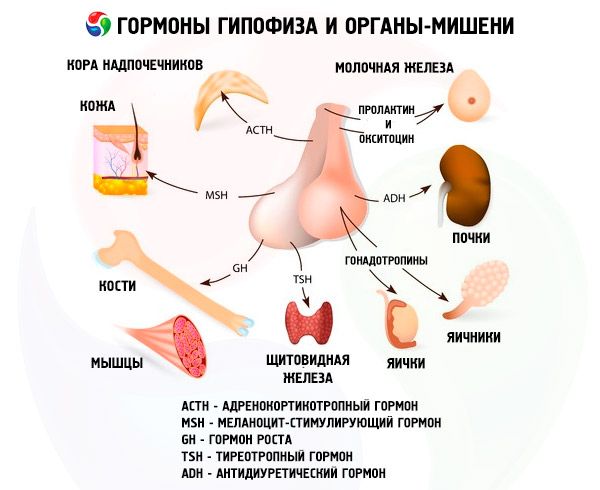
Hormoonidele eesmine ja tagumine ajuripatsis mõjutavad paljusid kehafunktsioonidele, eriti läbi teiste endokriinnäärmete. Eesmises lobe ajuripatsi acidophilic adenocytes (alfa) rakud toodavad somotropny hormooni (HGH), osalevad regulatsioonis kasvu ja arengut noore organismi. Kortikotropnye endocrinocytes eritavad adrenokortikotroopne hormoon (ACTH), mis stimuleerib sekretsiooni steroidhormoonide neerupealised. Tirotropnye endocrinocytes eritavad tirotropny hormooni (TSH), mis mõjutab kilpnäärme arengu ja aktiveerides tootmiseks selle hormoonid. Gonadotroopsed hormoonid: folliikuleid stimuleeriv hormoon (FSH), luteiniseeriv hormoon (LH) ja prolaktiini - mõjutada puberteedi keha, reguleerivad ja stimuleerivad folliikulite arengut munasarjas, ovulatsioon, rinna- kasvu ja piimatootmise naistel, protsess spermatogeneesi meestel. Neid hormoone toodavad beetarakkude basofiilsed adenotsüüdid . Siin lipotropic tegurid, mida eritab ajuripats, mis mõjutavad varumise ja kasutamise rasva kehas. Vaheosa eesmise sagara moodustatud melanotsüüte stimuleeriv hormoon, mis reguleerib moodustumist pigmente - melaniini - organismis.
Hüpotalamuse supraoptiliste ja paraventrikulaarsete tuumade neurosekretoorsed rakud toodavad vasopressiini ja oksütotsiini. Need hormoonid transporditakse Neurohüpofüüs rakud piki aksonit lahutamatuks Hüpotalaamuse-hüpofüseaalse põletikud. Hüpofüüsi tagajärjel esinevad need ained verest. Hormooni vasopressiini on veresooni ahendav ja antidiureetilise tegevus, mille eest ta sai nime antidiureetilise hormooni (ADH). Oksütotsiin on stimuleeriv mõju kontraktiivsus lihased emakas, suurendab piimas imetavate rinnanäärmed, pärsib arengut ja kollaskeha funktsiooni, mõjutab sile hääletooni muutmata (neischerchennyh) lihased seedetraktis.
Hüpofüüsi kujunemine
Hüpofüüsi esiosa areneb suu lahe seljaosa epiteelist ringikujulise väljakasvu kujul (Rathke tasku). See ektoparmaalne eend kasvab tulevase III ventrikli põhja poole. Teisele ajukoe põletikust (kolmanda vatsakese tulevane põhi) jõuab ta teise väljapoole, mille tagajärjel tekib lehtri teraline kraater ja ajuripatsi tagajalg.
 [1], [2], [3], [4], [5], [6], [7], [8], [9], [10], [11], [12], [13]
[1], [2], [3], [4], [5], [6], [7], [8], [9], [10], [11], [12], [13]
Hüveapõletiku kohad ja närvid
Ülemise ja alumise ajuripatsi arterid suunatakse ajuarteri ringi sisekordiarteritest ja veresoontesse hüpofüüsi. Ülemised hüpofüüsiarterid lähevad hüpotalamuse halli tuumale ja lehtrile, anastomoosid siin üksteisega ja moodustavad kapillaarid, esmane hemokapillaarne võrk, mis siseneb ajukudesse. Selle võrgu pikkadest ja lühikestest silmadest moodustuvad portaalveenid, mis on suunatud hüpofüüsi eesmisele ahvenale. Hüpofüüsi esiosa parenüühimas on need veenid jagatud laiade sinusoidaalseteks kapillaarideks, mis moodustavad sekundaarse hemokapillaarse võrgu. Hüpofüüsi tagaosas on peamiselt veri, mis voolab läbi madalama hüpofüüsiartikli. Ülemise ja alumise ajuripatsi arterite vahel on pikad arteriaalsed anastomoosid. Sekundaarse hemokapillaarvõrgu venoosse vere väljavoolu teostab veenide süsteem, mis voolab aju kõva kestani kõhukinnisesse ja piklikesse ninaotsadesse.
Hüpofüüsi innervatsioon sisaldab sümpaatilisi kiude, mis läbivad elundi koos arteritega. Postganglionaalsed sümpaatilised närvikiud liiguvad sisemise unearteri kokkutõmbumisest. Peale selle on hüpofüüsi tagajalal leitud hulgaliselt hüpotalamuse tuumades paiknevate neuronekretoorsete rakkude väljakasvu.
Hüpofüüsi vanuse omadused
Hüpofüüsi keskmine kaal vastsündinutel ulatub 0,12 g-ni. Kehakaal kahekordistab 10 ja kolmekordistub 15 aasta võrra. 20 aasta vanuseni jõuab hüpofüüsi mass maksimumini (530-560 mg) ja järgnevatel vanustel ei muutu peaaegu. 60 aasta pärast on selle sisesekretsiooni näärme mass vähenenud.
[14], [15], [16], [17], [18], [19], [20], [21], [22], [23], [24], [25], [26]
Hüpofüüsihormoonid
Närvisüsteemi ja hormonaalse regulatsiooni ühtsus organismis tagatakse hüpofüüsi ja hüpotalamuse lähedase anatoomilise ja funktsionaalse ühendusega. See kompleks määrab kogu sisesekretsioonisüsteemi seisundi ja toimimise.
Peamiseks Umpirauhanen mis toodab mitmeid peptiidhormoonide mis otseselt reguleerida funktsioonina perifeerse näärmed - ajuripatsis. On punaka värvusega oa-kujulise, hõlmatud kiulise kapsli kaaluga 0,5-0,6 g See veidi erinev sõltuvalt soost ja vanusest inimene. On üldtunnustatud jagunemise ajuripatsis kaheks osaks, erinevaid areng, struktuuri ja funktsiooni: esi- distaalse - hüpofüüsi eessagara ja tagasi - neurohypophysis. Esimese umbes 70% kogumassist eesnäärme ja jaguneb distaalne, Voronkov ja vaheosa, teine - tagumisel osal või fraktsioonis ja ajuripatsi vars. Võlli paikneb ajuripatsis lohk sella kiilukujulise luu ja läbi jala on ühendatud aju. Ülemine osa on hõlmatud anterior lobe Nägemisristmik ja nägemistrakti. Perfusiooni ajuripatsi väga rikkalikult oksad ja viiakse läbi sisemise unearteri (ülemise ja alumise ajuripatsi arterites) ja harude ajuarteri ringi. Ülem hüpofüseaalse arteri verevarustuse kaasatud adenohypophysis ja alumine - neurohypophysis mille puhul kontakt koos neurosekretoorsete aksoni lõpud hüpotalamuse magnocellular. Esimene osa mediaan ülekaaluga hüpotaalamuse mis on hajutatud kapillaaris võrgu (primaarne kapillaaride põimiku). Need kapillaarid (mis kontaktklemme aksonite väikeste mediobasaalsetes hüpotalamuse neurosekretoorsete rakud) kogutud värativeeni laskumisel mööda jalgade ajuripatsis adenohypophysis parenhüümi vajaduse jaotatakse jälle sinusoidi kapillaarid ahela (sekundaarne kapillaaride põimiku). Seega vere eelnevalt läbi mediaan ülekaaluga hüpotalamuse kus adenogipofizotropnymi rikastatud hüpotalamuse hormooni (kortikotropiini vabastav hormoon), jõuab adenohypophysis.
Väljavool veri, küllastunud adenogipofizarnymi hormoonid arvukate kapillaarid sekundaarse põimiku veenid viiakse läbi süsteemi, mis omakorda suubub venoosse ninakõrvalkoobaste kohta kõvakesta ja vereringesse. Seega on hüpotalamuse verevoolu hüpofüüsi portaal süsteem, mis on adenohüpofüüsi troofiliste funktsioonide neurohumoraalse kontrolli funktsioonide kompleksse mehhanismi morfofunktsionaalne komponent.
Hüpofüüsi inervatsiooni teostavad sümpaatilised kiud, mis järgivad hüpofüüsi artereid. Alustades neile antakse postganglioni kiude, läbides sisemist karotiidipelki, mis on ühendatud ülemiste emakakaela sõlmedega. Hüpotalamust ei toimu adenohüpofüüsi otsene inervatsioon. Hüpotalamuse neurosekretoorsete tuumade närvikiud sisenevad tagumisse serva.
Adenohüpofüüs histoloogilises arhitektoonikas on väga keeruline moodus. See eristab kahte liiki näärmetevahelisi rakke - kromofoobilist ja kromofiilset. Need omakorda jagunevad eosinofiilide ja basofiili (ajuripatsi üksikasjaliku histoloogilise kirjeldus on toodud vastavas osas käsitsi). Siiski tuleb märkida, et hormoonid toodetud näärmeliste rakud, mis moodustavad parenhüümi adenohypophysis tõttu viimastel mitmekesisuse mõnevõrra erinev nende keemilist olemust ja peenstruktuuri sekretiziruyuschih rakkude peaks vastama biosünteesi omadused iga. Kuid adenohüpofüüsi mõnikord on võimalik jälgida näärme-rakkude üleminekuvorme, mis on võimelised tekitama mitmeid hormoone. On tõendeid selle kohta, et adenohüpofüüsi mitmesuguseid näärme-rakke ei määratleta alati geneetiliselt.
Türgi saduli diafragma all on eesmise laba lehtri osa. See katab hüpofüüsi jalga, mis puutub halli küngasesse. Seda adenohüpofüüsi osa iseloomustab epiteelirakkude olemasolu ja rohkearvuline verevarustus. See on ka hormoonide aktiivsus.
Hüpofüüsi keskmine (keskmine) osa koosneb mitmest suure sekretsioon-aktiivse basofiilsete rakkude kihist.
Hüpofüüs oma hormoonide kaudu täidab erinevaid funktsioone. Oma anterior lobe toodetud adrenokortikotropiini (ACTH), kilpnääret stimuleeriv (TSH), folliikuleid stimuleeriv hormoon (FSH), luteiniseeriv hormoon (LH), lipotropic hormoon ja kasvuhormooni -. Somatotroopsed (SRT ja prolaktiini kesksagara sünteesiti melanotsüüte stimuleeriva hormooni (MSH), ja Seljaosas koguneb vasopressiin ja oksütotsiin.
AKGG
Hüpofüüsihormoonid esindavad valkude ja peptiidhormoonide ja glükoproteiinide rühma. Hüpofüüsi esiosa hormoonidest ACTH on kõige uuritud. Seda toodavad basofiilrakud. Selle peamine füsioloogiline funktsioon on biosünteesi stimulatsioon ja steroidhormoonide sekretsioon neerupealiste koorega. AKTHil on ka melanotsüütide stimuleeriv ja lipotroopne aktiivsus. Aastal 1953 eraldati see puhtal kujul. Hiljem loodi tema keemiline struktuur, mis koosnes 39 aminohappejäägist inimest ja mitmest imetajast. ACTH-l puudub spetsiifiline spetsiifilisus. Praegu viiakse läbi nii oma hormooni kui ka mitmesuguste, aktiivsemate kui looduslike hormoonide keemiline süntees, selle molekuli fragmendid. Hormooni struktuuris on peptiidiahela kaks sektsiooni, millest üks sisaldab ACTH tuvastamist ja seondumist retseptoriga, teine - annab bioloogilise efekti. ACTH retseptoriga tundub, et see seostub hormooni ja retseptori elektriliste laengute vastastiktoimetest. Bioloogilise efektori ACTH roll annab molekuli 4-10 fragmendi (Met-Glu-Gis-Fen-Arg-Tri-Tri).
ACTH-i melanotsüütide stimuleeriv aktiivsus tuleneb N-terminaalse piirkonna molekulis, mis koosneb 13 aminohappejäägist ja a-melanotsüütide stimuleeriva hormooni struktuuri kordamisest. Sama koht sisaldab heptapeptiidi, mis esineb teistes hüpofüüsi hormoonides ja millel on mõni adrenokortikotroopne, melanotsüüte stimuleeriv ja lipotroopne toime.
Võtmepunktiks toimel ACTH tuleks pidada aktiveerimist ensüümvalgu tsütoplasmas cAMP. Fosforüülitud proteiinkinaasi aktiveerib ensüümi Esteraasi muundab kolesterooliestrid vabastamiseks rasvainet tilkades. Valk sünteesitakse tsütoplasmas tulemusena fosforüülimise ribosoomiseondumise vabast kolesteroolist stimuleerib tsütokroom P-450 ja viia see lipiiditilku mitokondrites, kus on kõik ensüümid tagada kolesterooli muundumist kortikosteroide.
[27], [28], [29], [30], [31], [32], [33], [34], [35], [36], [37], [38], [39]
Tüotroopne hormoon
TSH - türeotropiin - kilpnäärme arengu ja funktsiooni peamine regulaator, kilpnäärmehormoonide sünteesi ja sekretsiooni protsessid. See kompleks valk - glükoproteiin - koosneb alfa- ja beeta-subühikutest. Esimese alaühiku struktuur langeb kokku luteiniseeriva hormooni alfa-alamühikuga. Pealegi langeb see suures osas erinevate loomaliikide hulka. Inimese TSH inimese beeta-subühiku aminohappejääkide järjestus detekteeritakse ja koosneb 119 aminohappejäägist. Võib märkida, et inimese TSH ja veise beeta-subühikud on paljudes aspektides sarnased. Glükoproteiini hormoonide bioloogilise aktiivsuse bioloogilised omadused ja iseloom määrab kindlaks beeta-alaühiku. Samuti tagab see hormooni koostoime retseptoritega erinevates sihtorganites. Kuid beeta subühik enamikus loomades näitab spetsiifilist aktiivsust alles pärast selle ühendamist alfa-subühikuga, mis toimib hormooni aktivaatorina. Viimane põhjustab sama tõenäosusega β-alaühiku omadustega määratud luteiniseerivat, folliikuleid stimuleerivat ja türeotroopset aktiivsust. Leitud sarnasus lubab meil järeldada, et need hormoonid pärinevad evolutsiooni käigus ühest lähteainest, beeta-subühik määrab hormoonide immunoloogilised omadused. Seal on oletus, et alfa-subühiku kaitseb beeta-subühik proteolüütiliste ensüümide toimel, ning samuti lihtsustab selle transportimist ajuripatsis perifeerse organam- "sihtmärgid".
Gonadotropilised hormoonid
Gonadotropiinid esinevad kehas LH ja FSH kujul. Nende hormoonide funktsionaalne eesmärk vähendab tavaliselt sugupoolteprotsesside pakkumist mõlema soo indiviidides. Need, nagu TTG, on komplekssed valgud - glükoproteiinid. FSH indutseerib naistel munarakkude folliikulite küpsemist ja stimuleerib spermatogeneesi meestel. LH põhjustab naiste folliikulite rebenemist kollase keha moodustumisega ja stimuleerib östrogeenide ja progesterooni sekretsiooni. Meestel kiirendab see sama hormoon interstitsiaalse kude ja androgeenide sekretsiooni. Gonadotropiinide toime sõltub üksteisest ja jätkub sünkroonselt.
Gonadotropiini sekretsiooni dünaamika naistel erineb menstruaaltsükli ajal ja seda uuritakse piisavalt üksikasjalikult. Tsükli preovulatoorses (follikulaarses) faasis on LH sisaldus üsna madal ja FSH on suurenenud. Kuna folliikulite küpsemise sekretsioon östradiooli suurenenud, suurendades seeläbi tootmise Gonadotropiinide ajuripatsi poolt ja välimus tsüklit nii LH kui ka FSH nii edasi. E., suguhormoonid stimuleerivad gonadotropiinidele.
Praegu määratakse LH struktuur. Nagu TTG, koosneb see 2 allüksusest: a ja p. LH alfa-alaühiku struktuur erinevatel loomaliikidel langeb suures osas kokku, see vastab TSH alfa-subühiku struktuurile.
Struktuuri beetasubühiku LH erineb oluliselt struktuuri TSH beeta-subühiku, kuigi see on neli võrdne kogus peptiidahela koosseisus 4-5 aminohappejääki. TTG-s paiknevad nad positsioonides 27-31, 51-54, 65-68 ja 78-83. Kuna beeta-subühiku LH ja TSH määrab spetsiifiline bioloogiline aktiivsus hormoonid, siis võib eeldada, et homoloogsete piirkondade struktuuri LH ja TSH peaks andma beetasubühiku alfa-subühiku ja erinevad oma struktuurilt krunti - vastutavad spetsiifilisust bioloogilist aktiivsust hormoon.
Native LH on väga stabiilsed proteolüütiliste ensüümide toimel, kuid beeta-subühiku kiiresti lõhustada kümotrüpsiin ja kõva-subühiku hüdrolüüsitakse ensüüm, st. E. See on kaitsev roll, mis takistab juurdepääsu kümotrüpsiinipeptiidi sidemetega.
Mis puudutab FSH keemilist struktuuri, siis praegu ei ole teadlased lõplikke tulemusi saavutanud. Sarnaselt LH-ga sisaldab FSH kahte subühikut, aga FSH beeta-subühik erineb LH beeta-subühikust.
Prolaktiin
Taastumisprotsessides osaleb aktiivselt teine hormoon, prolaktiin (laktootiline hormoon). Imetajatele prolaktiini peamised füsioloogilised omadused ilmnevad piimanäärmete ja imetamise, rinnakarglite ja siseorganite kasvu stimulatsiooni vormis. See soodustab steroidide mõju sekundaarsetele seksuaalomadustele meestel, stimuleerib kollase keha sekretoorset aktiivsust hiirtel ja rottidel ning osaleb rasvade metabolismi reguleerimises. Viimastel aastatel on prolaktiinist palju tähelepanu pööratud emade käitumisharjumuste regulatsioonile, seda polüfunktsionaalsust seletatakse selle evolutsioonilise arenguga. See on üks iidsetest hüpofüüsi hormoonidest ja seda leidub ka kahepaiksed. Praegu on mõne imetaja liigi prolaktiini struktuur täielikult detekteeritud. Kuid kuni viimase ajani on teadlased väljendanud kahtlusi sellise hormooni olemasolust inimestel. Paljud uskusid, et selle funktsiooni teostab kasvuhormoon. Nüüd on inimestel veenvad tõendid prolaktiini esinemise kohta ja selle struktuur osaliselt dešifreeritud. Prolaktiini retseptorid seonduvad aktiivselt kasvuhormooni ja platsenta lakto-geeniga, mis näitab kolme hormooni ühe toimemehhanismi.
Somatotropiin
Veelgi laiemast toimet kui prolaktiinil on kasvuhormoon - somatotropiin. Nagu prolaktiin, toodetakse seda adenohüpofüüsi acidophilic rakkudes. STG stimuleerib skeleti kasvu, aktiveerib valgu biosünteesi, annab rasva mobiliseeriva efekti, soodustab keha suuruse suurenemist. Lisaks koordineerib ta vahetusprotsesse.
Hormooni osalemine viimases on kinnitust leidnud hüpofüüsi sekretsiooni järsu suurenemise, näiteks vere suhkrusisalduse vähenemisega.
Selle inimese hormooni keemiline struktuur on nüüd täielikult välja kujunenud - 191 aminohappejääki. Selle esmane struktuur sarnaneb koorion-somatomamotropiini või platsenta lakto-geeni struktuuriga. Need andmed näitavad kahe hormooni olulist evolutsioonilist lähedust, kuigi neil on erinevusi bioloogilises aktiivsuses.
On vaja rõhutada kõnealuse hormooni kõrget spetsiifilisust - näiteks loomse päritoluga STH on inaktiivne inimestel. See on tingitud nii inimese ja loomade STH-retseptorite kui ka hormooni enda struktuurist. Praegu on käimas uuringud, et identifitseerida aktiivsetele kohtadele STH kompleksstruktuuri, millel on bioloogiline aktiivsus. Uurime molekuli individuaalseid fragmente, millel on muud omadused. Näiteks pärast inimese STH hüdrolüüsi pepsiiniga eraldati peptiid, mis sisaldas 14 aminohappejääki ja vastas molekuli 31-44 piirkonda. Tal ei olnud kasvu mõju, kuid lipotroopne aktiivsus oli oluliselt parem natiivse hormooniga. Inimese kasvuhormoon, erinevalt loomadel sarnastest hormoonidest, omab olulist laktootilist toimet.
Paljudel adenohypophysis sünteesida nii peptiidi ja valkained võttes rasva mobiliseeriv toime ja tropic hüpofüüsihormoonidele - ACTH, kasvuhormoon, TSH ja muud - kutsuda Lipotroopsena efekti. Viimastel aastatel on välja toodud eelkõige beeta- ja y-lipotroopsed hormoonid (LPG). Kõige laialdasemalt uuritud bioloogiliste omaduste beeta-LPG, mis lisaks lipotropic aktiivsusel on ka melanotsüüte, kortikotropinstimuliruyuschee ja hüpokaltseemiiliselt mõju ning see annab insuliini toime.
Praegu detekteeritakse lamba veeldatud naftagaasi (90 aminohappejäägi), sigade ja veiste lipotroopsete hormoonide peamist struktuuri. Sellel hormoonil on spetsiifiline spetsiifilisus, kuigi beeta-LPG keskosa osa struktuur erinevates liikides on sama. See määrab hormooni bioloogilised omadused. Selle ala üks fragmente on leitud alfa-MSH, beeta-MSH, ACTH ja beeta-LPG-i struktuuris. On soovitatav, et need hormoonid pärinevad evolutsiooni käigus samalt prekursorilt. Y-LPG-l on vedela õli pehmem lipotroopne aktiivsus.
Melanotsüütide stimuleeriv hormoon
See hormoon sünteesitakse kesksagara ajuripatsi, tema bioloogilise funktsiooni stimuleeriva naha pigmendi melaniini biosünteesi, suurendab suurus ja kogus pigmendiga melanotsüüdid rakud kahepaiksed. Neid MSH omadusi kasutatakse hormooni bioloogilises testis. On kahte tüüpi hormoone: alfa ja beeta-MSH. On näidatud, et alfa-MSH-il pole spetsiifilist spetsiifilisust ja kõigil imetajatel on sama keemiline struktuur. Selle molekuliks on peptiidiahel, mis koosneb 13 aminohappejäägist. Seevastu beeta-MSH omab spetsiifilist spetsiifilisust ja selle struktuur erineb loomadel. Enamikel imetajatel on β-MSH molekul koosnevad 18 aminohappejäägist ja inimestel on see aminotsadest piklik kuni neli aminohappejääki. Tuleb märkida, et alfa-MSH-il on mõni adrenokortikotroopne aktiivsus ning selle mõju loomade ja inimeste käitumisele on nüüd tõestatud.
Oksütotsiin ja vasopressiin
Tagumisse lobe ajuripatsi koguneda vasopressiini ja oksütotsiini, mis on sünteesitud hüpotalamuse: vasopressiini - neuronites kohta supraoptic tuumas ja oksütotsiini - paraventrikulyatornogo. Siis viiakse nad hüpofüüsi. Tuleb rõhutada, et hüpotalamuses sünteesitakse algul eellase vasopressiini hormoon. Samas tekib seal 1. Ja 2. Tüüpi neurofisiini valk. Esimene seob oksütotsiini ja teine vasopressiini. Need kompleksid migreeruvad neurosekretoorsete graanulid tsütoplasmas piki aksonit ja jõuab Neurohüpofüüs kus närvikiude lõppeda veresoone seina ja graanulite verre. Vasopressiini ja oksütotsiini - esimese hüpofüüsihormoonidele täis aminohappe järjestus. Nende keemilises struktuuris on nad ühe disulfiidsillaga nonapeptiidid.
Peetakse hormoonid toota erinevaid bioloogilisi toimeid: stimuleerida transpordi kloriidi ja vee läbi membraani, on pressortoimest, suurendab kontraktsiooni silelihaste emaka sünnituse ajal, suurendab sekretsiooni piimanäärmetes. Tuleb märkida, et vasopressiinil on antidiureetiline toime kõrgem kui oksütotsiini, samas kui viimane toimib tugevamalt emakas ja rinnanäärmes. Peamiseks regulaatorina vasopressiini eritumise on veetarbimine neerutuubulitesse seondub see retseptorite tsütoplasmamembraan ja sellele järgnev aktiveerimine ensüümi adenülaattsüklaasi neid. Hormooni sidumiseks retseptoriga ja bioloogilise efektiga vastutavad molekuli erinevad osad.
Hüpofüüsi seostatakse hüpotalamuse kaudu närvisüsteemi, ühendab täisarv funktsionaalse endokriinsüsteemi kaasatud tagamisel püsivuse sisekeskkonna (homöostaasi). Sees endokriinsed homeostatic määrus põhineb põhimõttel tagasisidet vahel anterior lobe ajuripatsi ja zhelezami- "sihtmärgid" (kilpnääre, neerupealise koore, sugunäärmete). Liigne hormoon, mida toodetakse zhelezoy- "sihtmärk", aeglane ning selle puuduse stimuleerib sekretsiooni ja eraldamist vastavatest tropic hormoon. Tagasiside süsteem sisaldab hüpotalamust. See on selles, et need on tundlikud raua sihtmärkide, retseptori tsoonide hormoonide suhtes. Spetsiifiliselt seostuma ringlevast hormoonid veres ja muutuvate vastuse sõltuvalt kontsentratsioonist hormoonid, hüpotalamuse retseptorite edastab selle mõju asjakohasele hüpotalamuse keskuste et töö koordineerimiseks hüpofüüsi eessagaras, hüpotalamuse hormoone vabastavate adenogipofizotropnye. Seega peaks hüpotalamust pidama neuro-endokriinseks ajjuks.