
Katehhoolamiinide süntees, sekretsioon ja ainevahetus
Viimati vaadatud: 19.10.2021

Kõik iLive'i sisu vaadatakse meditsiiniliselt läbi või seda kontrollitakse, et tagada võimalikult suur faktiline täpsus.
Meil on ranged allhanke juhised ja link ainult mainekate meediakanalite, akadeemiliste teadusasutuste ja võimaluse korral meditsiiniliselt vastastikuste eksperthinnangutega. Pange tähele, et sulgudes ([1], [2] jne) olevad numbrid on nende uuringute linkideks.
Kui tunnete, et mõni meie sisu on ebatäpne, aegunud või muul viisil küsitav, valige see ja vajutage Ctrl + Enter.
Neerupealiste ajuümbris moodustab struktuuri steroidide kaugel olevast ühendist. Need sisaldavad 3,4-dihüdroksüfenüül (katehhool) tuuma ja neid nimetatakse katehhoolamiinideks. Nende hulka kuuluvad adrenaliin, norepinefriin ja dopamiin beeta-oksütiramiin.
Katehhoolamiinide sünteesi järjestus on üsna lihtne: türosiin → dioksüfenüülalaniin (DOPA) → dopamiin → noradrenaliin → adrenaliin. Türosiin siseneb kehasse toiduga, kuid see võib moodustuda ka fenüülalaniinist maksa fenüülalaniini hüdroksülaasi toimel. Türosiini transformatsiooni lõpptooted kudedes on erinevad. Neerupealise säsi protsess jätkub etapiga adrenaliin moodustumise otstes sümpaatilise närvi - noradrenaliini teatud neuronite kesknärvisüsteemi katehhoolamiinide dopamiini sünteesi täidetud vormi.
Türosiini konverteerimine DOPA-sse katalüüsitakse türosiini hüdroksülaasiga, millest koosseisus on tetrahüdro-biopteriin ja hapnik. Usutakse, et see ensüüm piirab kogu katehhoolamiini biosünteesi protsessi kiirust ja protsessi lõpptooted neid inhibeerivad. Katehhoolamiinide biosünteesi regulatiivsete mõjude põhieesmärk on türosiini hüdroksülaas.
Dopa dopamiini konversioon mida katalüüsib ensüüm dopa dekarboksülaasi (kofaktor - pyridoxal), mis on suhteliselt mittespetsiifilised ja dekarboksüleerida ja teiste aromaatsete L-aminohappe. Siiski on märke selle kohta, et katehhoolamiine võib sünteesida, muutes aktiivsust ja seda ensüümi. Mõnes neuronis ei ole dopamiini edasiseks muundamiseks ensüüme, see on lõpptooteks. Teised kuded sisaldavad dopamiin-beeta-hüdroksülaasi (kofaktorid - vask, askorbiinhape ja hapnik), mis muudab dopamiini norepinefriiniks. Neerupealise säsi (kuid mitte sümpaatiline närvilõpmeid) esineb fenüületanoolamiin- - metüültransferraasist moodustav adrenaliin alates noradrenaliini. Metüülrühmade doonor on sel juhul S-adenosüülmetioniin.
Oluline on meeles pidada, et sünteesi fenüületanoolamiin--N-Metiltransferazy indutseeritud glükokortikoidide langevad Ajukoore kihi värativeenisüsteem. See võib seletada asjaolu, valesid Kombineerides kahte erinevat endokriinnäärmetes ühes kehas. Mõiste glükokortikoidi sünteesi adrenaliin rõhutas ka asjaolu, et neerupealise säsi rakud, mis toodavad norepinefriini, paigutatud ümber arteriaalse laevadel ning vererakkude Saadakse adrenalinprodutsiruyuschie sisuliselt venoosse ninakõrvalkoopad lokaliseeritud neerupealise koores.
Kokkuvarisemist katehhoolamiinide esineb peamiselt mõjul kahe ensüümi süsteemid: katehhoolvaheastmete-Ometüültransferaasi (COMT) ja (MAO). Peamist võimalust epinefriini ja norepinefriini kõdunemise skemaatiliselt kujutatud joonisel. 54. Toimel COMT kohalolekul doonoriks metüülrühmad S-adrenozilmetionina katehhoolamiinide ja muundatakse normetanefriini metanefriini (3-O-metüül-derivaatide epinefriini ja norepinefriini), mille mõjul MAO muundatakse aldehüüdide ja rohkem (juuresolekul aldehüüd) lahust vanillüül-mandelhape hape (ICH) - põhiline lagusaadus noradrenaliini ja adrenaliini. Samal juhul, kui esimene kokkupuude katehhoolamiinide MAO tegevus, mitte COMT neid muundada 3,4-dioksimindalevy aldehüüdi ja seejärel mõjul aldehüüdi ja COMT - 3,4-dioksimindalnuyu happe ja ESV. Kohalolekul alkoholdehüdrogenaas katehhoolamiinide võivad moodustada 3-metoksü-4-oksifenilglikol peamises lõpp-produktiks lagunemise epinefriini ja norepinefriini kesknärvisüsteemis.
Desintegreerumiskatse dopamiini kulgeb sarnaselt, va selle metaboliitide puudub hüdroksüülrühmade beeta-süsinikuaatom, mistõttu asemel vanillüül-mandelhape moodustatud homovanilliinhape (HVA) ja 3-metoksü-4-oksifeniluksusnaya happega.
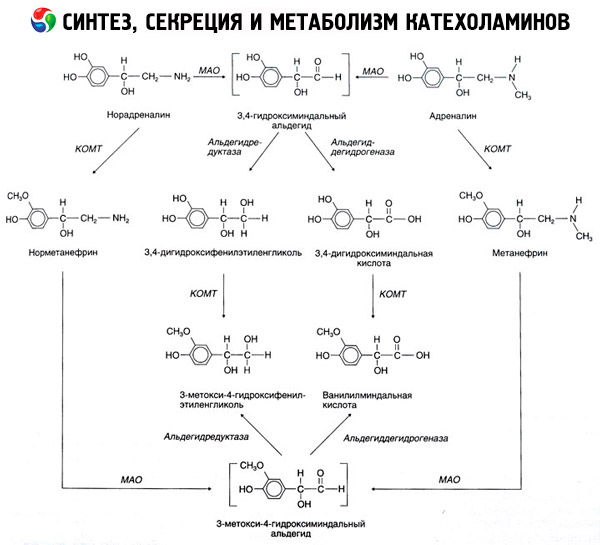
Positiivseks võib olla ka kino-aminorühma molekuli oksüdeerumisega seotud kinoidrajati olemasolu, millel võib olla väljendunud bioloogilise aktiivsusega vaheprodukte.
Moodustati toimel tsütosoolensüümi, adrenaliin ja noradrenaliin sümpaatilise närvilõpmeid, neerupealise säsi ja sisesta sekretoorse graanulid, mis kaitseb neid toime degradatsiooniensüümid. Katehhoolamiinide püüdmine graanulitega nõuab energiakulusid. In chromaffin graanulid neerupealise säsi katehhoolamiinide kindlalt seotud ATP (vahekorras 4: 1) ja spetsiifilisi valke - Kromograniin mis takistab difusiooni pärit hormoonide graanulid tsütoplasmas.
Otsene stiimuli sekretsiooni katehhoolamiinide nähtavasti penetratsiooni kaltsiumi raku stimuleeriva eksotsütoosiga (fusion membraani graanulite rakupinnal ja nende mahajäämus üldsaagises lahustuva vormi sisaldus - katehhoolamiinide, dopamiini-beeta-hüdroksülaasi ATP ja Kromograniin - rakuvälisesse vedelikku) .
Katehhoolamiinide füsioloogilised mõjud ja nende toimemehhanism
Katehhoolamiinide toime algab koostoimel sihtrakkude spetsiifiliste retseptoritega. Kui kilpnäärme- ja steroidhormoonide retseptorid paiknevad rakkude sees, paiknevad katehhoolamiini retseptorid (samuti atsetüülkoliini ja peptiidhormoonid) välimise raku pinnal.
See on juba ammu kindlaks tehtud, et seoses mõned reaktsioonid adrenaliin või noradrenaliin on tõhusamad kui sünteetilised katehhoolamiinide isoproterenoolile, teiste efekt on parem toime isoproterenoolile epinefriini või norepinefriini. Sellest lähtuvalt töötati välja idee kahe tüüpi adrenoretseptorite esinemise kohta kudedes: alfa ja beeta, mõnel neist võib esineda ainult üks neist kahest tüübist. Isoproterenooli on kõige tugevam agonisti beeta-adrenergiliste retseptorite, arvestades sünteetiline ühend fenülefriini - kõige tugevam agonisti alfa-adrenergiliste retseptorite suhtes. Looduslikud katehhoolamiinide - adrenaliin ja noradrenaliin - on võimelised suhtlema retseptoritele mõlemat tüüpi, kuid adrenaliin võtab suurem afiinsus beeta ja norepinefriini - alfa-retseptorite suhtes.
Katehhoolamiinide tugevamaks aktiveerimiseks südame beeta-adrenergiliste retseptorite kui beeta retseptoreid silelihaste, võimaldades beeta-tüüpi jagatud alatüüpide: beta1-retseptorid (süda, rasvarakkude) ja p2 retseptorid (bronhid, veresooned, jne ...). Toimel isoproterenoolile kohta beta1 retseptori laitmatu toime adrenaliin ja noradrenaliin ainult 10 korda, samal ajal kui beta2-retseptorite toimib 100-1000 korda kangem looduslikud katehhoolamiinide.
Spetsiifiliste antagonistide (fentaalamiin ja fenoksübensamiin alfa ja propranolooli kasutamine beeta retseptorite puhul) kasutamine kinnitas adrenoretseptorite klassifikatsiooni adekvaatsust. Dopamiin on võimelised suhtlema nii alfa- ja beeta-retseptorid, kuid erinevates kudedes (aju, ajuripatsis laevad) leitud ja enda dopamiinergiliste retseptorite spetsiifiliste blokeerija mis haloperidool. Beeta retseptorite arv vahemikus 1000 kuni 2000 rakku. Beeta retseptorite poolt vahendatud katehhoolamiinide bioloogilised mõjud on tavaliselt seotud adenülaattsüklaasi aktiveerimisega ja cAMP rakusisese sisalduse suurenemisega. Retseptor ja ensüüm, kuigi nad on funktsionaalselt ühendatud, kuid esindavad erinevaid makromolekule. Adenülaattsüklaasi aktiivsuse modulatsioonis on hormooni retseptori kompleksi, guanosiintrifosfaadi (GTP) ja teiste puriinnukleotiidide mõju kaasatud. Ensüümi aktiivsuse suurenemisega tundub, et need vähendavad agonistide beeta retseptorite afiinsust.
Detsentreeritud struktuuride tundlikkuse suurendamise nähtus on juba ammu teada. Vastupidi, pikaajaline kokkupuude agonistidega vähendab sihtkudede tundlikkust. Peptaarsete retseptorite uuring lubas neid nähtusi seletada. Näidati, et isoproterenooli pikaajaline toime põhjustab adenülaattsüklaasi tundlikkuse kaotuse, kuna beeta retseptorite arv väheneb.
Desensibiliseerimise protsess ei nõua valkude sünteesi aktiveerimist ja on tõenäoliselt tingitud pöördumatute hormooni-retseptori komplekside järk-järgulise moodustumisest. Vastupidi, 6-oksidofamiini manustamist, mis lõhub sümpaatilised otsad, kaasneb reageerivate beeta retseptorite arvu suurenemine kudedes. Ei ole välistatud, et sümpateetiliste närvisüsteemi aktiivsuse suurenemine määrab katehhoolamiinidega seotud veresoonte ja rasvkoe vanusest sõltuva desensibiliseerimise.
Erinevate elundite adrenoretseptorite arvu saab kontrollida teiste hormoonide poolt. Seega suurendab östradiooli, progesterooni ja arvu vähendamiseks alfa-adrenoretseptorite emakas, mis kaasneb vastav suurenemine ja vähenemine selle kontraktiilsusreaktsiooni katehhoolamiinide. Kui β-retseptori agonistide toimel moodustunud intratsellulaarne "teine messenger" on kindlasti cAMP, siis on alfa-adrenergiliste mõjude saatja olukord keerulisem. Eeldatakse, et on olemas mitmesugused mehhanismid: cAMP taseme langus, cAMP sisalduse suurenemine, kaltsiumi raku dünaamika modulatsioon,
Erinevate toimete paljunemiseks organismis on tavaliselt adrenaliini doosid, mis on 5 ... 10 korda madalamad kui noradrenaliini. Kuigi viimane on efektiivsem suhtes a- ja beta1-adrenergiliste retseptorite, on oluline meeles pidada, et nii endogeensed katehhoolamiinide võimelised suhtlema nii alfa- ja beeta-retseptoreid. Seepärast sõltub selle keha bioloogiline reaktsioon adrenergilise aktiveerimisega suuresti selles sisalduvatest retseptoritüüpidest. See aga ei tähenda, et sümpaatilise ja neerupealise närvisüsteemi või humoraalse seose selektiivne aktiveerimine on võimatu. Enamikul juhtudel on selle erinevate linkide intensiivne tegevus. Seega eeldatakse, et see aktiveerib refleksi hüpoglükeemia neerupealise säsi, samas kui vererõhu langus (posturaalne hüpotensioon), millega kaasneb peamiselt norepinefriini vabanemist sümpaatiline närvilõpmeid.
Adrenoretseptorid ja nende aktiveerumise mõjud erinevates kudedes
|
Süsteem, orel |
Adrenoceptoritüüp |
Reaktsioon |
|
Kardiovaskulaarsüsteem: |
||
|
Süda |
Beeta |
Kontraktsioonide sageduse, juhtivuse ja kontraktiilsuse suurenemine |
|
Arterioolid: |
||
|
Nahk ja limaskestad |
Alfa |
Vähendamine |
|
Skeletilihaseid |
Beeta |
Pikenduse vähendamine |
|
Kõhuorganid |
Alfa (rohkem) |
Vähendamine |
|
Beeta |
Pikendamine |
|
|
Veenid |
Alfa |
Vähendamine |
|
Hingamise süsteem: |
||
|
Bronhide lihased |
Beeta |
Pikendamine |
|
Seedetrakt: |
||
|
Kõhuga |
Beeta |
Motoorse funktsiooni vähenemine |
|
Soolestik |
Alfa |
Sphintsteri vähendamine |
|
Põrna |
Alfa |
Vähendamine |
|
Beeta |
Lõõgastumine |
|
|
Kõhunäärme välimine salajane osa |
Alfa |
Sekretsiooni vähenemine |
|
Kuseteede süsteem: |
Alfa |
Sphincter vähendamine |
|
Kusepõis |
Beeta |
Lõõgastav exorcist lihas |
|
Mehed, suguelundid |
Alfa |
Ejakulatsioon |
|
Silmad |
Alfa |
Õpilane laienes |
|
Nahk |
Alfa |
Suurenenud higistamine |
|
Süljenäärmed |
Alfa |
Kaaliumi ja vee eraldamine |
|
Beeta |
Amülaasi sekretsioon |
|
|
Endokriinsed näärmed: |
||
|
Kõhunääre saared |
||
|
Beeta-rakud |
Alfa (rohkem) |
Vähendatud insuliini sekretsioon |
|
Beeta |
Suurenenud insuliini sekretsioon |
|
|
Alfa-rakud |
Beeta |
Suurenenud glükagooni sekretsioon |
|
8-rakud |
Beeta |
Suurenenud somatostatiini sekretsioon |
|
Hüpotalamuse ja ajuripatsi: |
||
|
Somatotroofid |
Alfa |
STH suurenenud sekretsioon |
|
Beeta |
STH vähenenud sekretsioon |
|
|
Laktotroofid |
Alfa |
Prolaktiini sekretsiooni vähenemine |
|
Türeotroofid |
Alfa |
TSH sekretsiooni vähenemine |
|
Kortikotroofid |
Alfa |
ACTH suurenenud sekretsioon |
| beeta | ACTH sekretsiooni vähenemine | |
|
Kilpnääre: |
||
|
Follikulaarrakud |
Alfa |
Türoksiini sekretsiooni vähenemine |
|
Beeta |
Tiroktiini suurenenud sekretsioon |
|
|
Parafollikulaarse (K) rakud |
Beeta |
Kaltsitoniini suurenenud sekretsioon |
|
Paratüroidnäärmed |
Beeta |
PTH suurenenud sekretsioon |
|
Neerud |
Beeta |
Reniini sekretsiooni suurenemine |
|
Kõhu |
Beeta |
Suurendage gastriini sekretsiooni |
|
Põhjavahetus |
Beeta |
Hapniku tarbimise suurenemine |
|
Maks |
? |
Glükogenolüüsi ja glükoneogeneesi suurenemine glükoosi saagist; ketoonisisalduse suurendamine ketoonikoguste vabanemisega |
|
Rasvkude |
Beeta |
Libolüüsi suurenemine vabade rasvhapete ja glütserooli vabastamisega |
|
Skeletilihased |
Beeta |
Glükolüüsi suurenemine püruvaadi ja laktaadi vabanemisega; proteolüüsi langus alaniini, glutamiini saagise vähenemisega |
Oluline on meeles pidada, et tulemused intravenoosse manustamise katehhoolamiinide ei ole alati piisavalt kajastada mõju endogeense ühendeid. See kehtib peamiselt noradrenaliini, sest see seisab keha enamasti ei verd, kuid otse sünapsipilust. Seetõttu endogeensed norepinefriini aktiveerib näiteks mitte ainult vaskulaarse alfa retseptorid (kõrgenenud vererõhk), vaid ka südame beeta-retseptorid (südamepekslemine), arvestades manustamiseks noradrenaliini välise otsaga valdavalt selle versoone alfa retseptoriga ja refleksi (via vagus) aeglustamist südamepekslemine.
Epinefriini madalad annused aktiveerivad peamiselt lihaste veresoonte ja südame beeta retseptoreid, mille tulemuseks on perifeerse veresoonte resistentsuse vähenemine ja südame minutimahu suurenemine. Mõnedel juhtudel võib esmane efekt olla domineeriv ja pärast adrenaliini manustamist tekib hüpotensioon. Suuremate annuste korral aktiveerib adrenaliin alfa retseptoreid, millega kaasneb perifeerse vaskulaarse resistentsuse suurenemine ja südame minutimahu suurenemise taustal vererõhu tõus. Siiski säilib ka selle toime veresoonkonna beeta retseptoritele. Selle tulemusena ületab süstoolse rõhu tõus diastoolse rõhu sarnast väärtust (impulsi rõhu suurenemine). Suuremate annuste kasutuselevõtuga alustatakse domineerivate epinefriini alfa-mimeetilist toimet: süstoolne ja diastoolne rõhk suureneb paralleelselt nii noradrenaliini toimel.
Katehhoolamiinide toime ainevahetusele sõltub nende otsestest ja kaudsetest mõjudest. Esimesed teostatakse peamiselt beeta-retseptorite kaudu. Keerukamad protsessid on seotud maksaga. Kuigi maksa glükogenolüüsi suurendamist on traditsiooniliselt peetud beeta retseptori aktiveerimise tulemuseks, on olemas ka andmed alfa-retseptorite kaasamise kohta selles. Katehhoolamiinide vahendatud toimed on seotud paljude teiste hormoonide, näiteks insuliini sekretsiooni moduleerimisega. Sekretsiooni ajal adrenaliini toimel on selgelt ülekaalus alfa-adrenergiline komponent, kuna on näidatud, et mis tahes stressiga kaasneb insuliini sekretsiooni pärssimine.
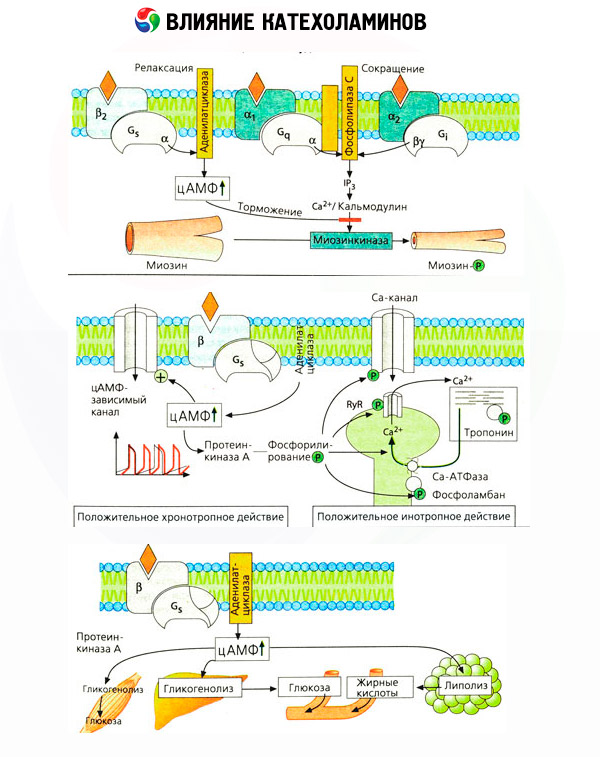
Katehhoolamiinide otsese ja kaudse toime kombinatsioon põhjustab hüperglükeemiat, mis on seotud mitte ainult glükoosi maksa suurenemisega, vaid ka selle kasutamise pärssimisega perifeersetes kudedes. Lipolüüsi kiirendamine põhjustab hüperlipatsideemiat, rasvhapete suuremat manustamist maksas ja ketooni kehade tootmise intensiivistamist. Glükolüüsi suurenemine lihastes suurendab laktaadi ja püruvaadi vabanemist veres, mis koos rasvkoest vabanenud glütserooliga toimib maksa glükoneogeneesi prekursoridena.
Katehhoolamiinide sekretsiooni reguleerimine. Sarnasus tooted ja meetodid vastuse sümpaatilise närvisüsteemi ja neerupealise säsi oli aluseks Kombineerides need struktuurid ühtseks tervikuks sympathoadrenal süsteemi vabanemisega neuraalne ja hormonaalne selle seos. Erinevad aferentsed signaalid on koondunud hüpotalamuse ja keskuste seljaaju ja piklikaju, millest pärinevad efferent maatüki sisselĂĽlitamine preganglionaarseid neuronrakkude asuvate organite külgmise sarve seljaaju tasemel emakakaela VIII - II-III nimme segmentides.
Preganglionaarseid aksonite nende rakkude lahkuvad seljaajust ja moodustada sünaptiline sidemed neuronid paiknevad ganglionides sümpaatilise ahelaga või neerupealise säsi rakud. Need preganglioni kiud on kolinergilised. Esimene põhiline erinevus postganglionic sümpaatiline neuroneid ja neerupealiste säsis kromafiinirakkudest seisneb selles, et viimane on edastatud sissetuleva signaali see koliinergilist neuro-juhtivuse (postganglionic adrenergilise närvid) ja humoraalne korostamisella adrenergilise ühendi veres. Teine erinevus on vähendatud postganglionic närve, mis toodavad norepinefriini, samas neerupealise säsi rakud - eelistatavalt adrenaliin. Need kaks ainet avaldavad kudele erinevat mõju.